

This is a preprint.
Visualization of translation reorganization upon persistent collision stress in mammalian cells
- PMID: 36993420
- PMCID: PMC10055323
- DOI: 10.1101/2023.03.23.533914
Visualization of translation reorganization upon persistent collision stress in mammalian cells
Update in
-
Visualization of translation reorganization upon persistent ribosome collision stress in mammalian cells.Mol Cell. 2024 Mar 21;84(6):1078-1089.e4. doi: 10.1016/j.molcel.2024.01.015. Epub 2024 Feb 9. Mol Cell. 2024. PMID: 38340715 Free PMC article.
Abstract
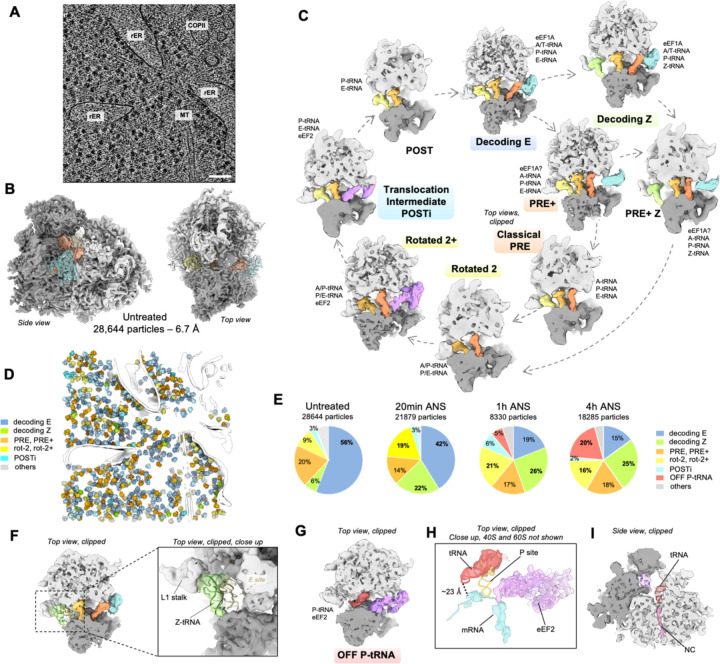
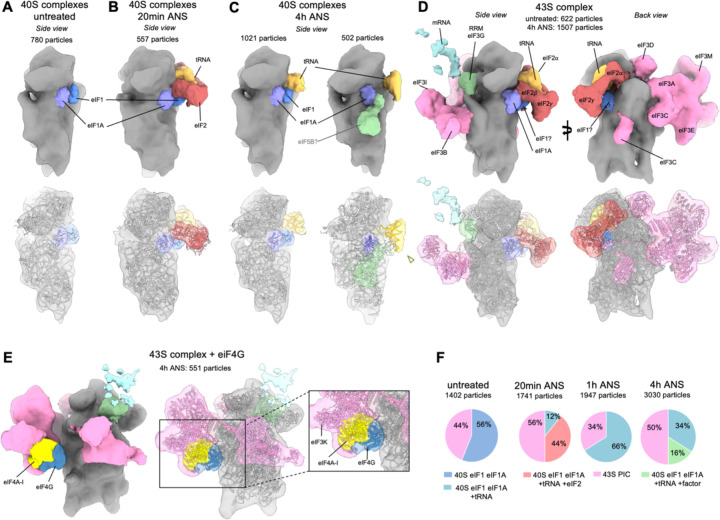
Aberrantly slow mRNA translation leads to ribosome stalling and subsequent collision with the trailing neighbor. Ribosome collisions have recently been shown to act as stress sensors in the cell, with the ability to trigger stress responses balancing survival and apoptotic cell-fate decisions depending on the stress level. However, we lack a molecular understanding of the reorganization of translation processes over time in mammalian cells exposed to an unresolved collision stress. Here we visualize the effect of a persistent collision stress on translation using in situ cryo electron tomography. We observe that low dose anisomycin collision stress leads to the stabilization of Z-site bound tRNA on elongating 80S ribosomes, as well as to the accumulation of an off-pathway 80S complex possibly resulting from collision splitting events. We visualize collided disomes in situ, occurring on compressed polysomes and revealing a stabilized geometry involving the Z-tRNA and L1 stalk on the stalled ribosome, and eEF2 bound to its collided rotated-2 neighbor. In addition, non-functional post-splitting 60S complexes accumulate in the stressed cells, indicating a limiting Ribosome associated Quality Control clearing rate. Finally, we observe the apparition of tRNA-bound aberrant 40S complexes shifting with the stress timepoint, suggesting a succession of different initiation inhibition mechanisms over time. Altogether, our work visualizes the changes of translation complexes under persistent collision stress in mammalian cells, indicating how perturbations in initiation, elongation and quality control processes contribute to an overall reduced protein synthesis.
Conflict of interest statement
Declaration of interest The authors declare no competing interest.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous