

This is a preprint.
Foundation model of neural activity predicts response to new stimulus types and anatomy
- PMID: 36993435
- PMCID: PMC10055288
- DOI: 10.1101/2023.03.21.533548
Foundation model of neural activity predicts response to new stimulus types and anatomy
Update in
-
Foundation model of neural activity predicts response to new stimulus types.Nature. 2025 Apr;640(8058):470-477. doi: 10.1038/s41586-025-08829-y. Epub 2025 Apr 9. Nature. 2025. PMID: 40205215 Free PMC article.
Abstract
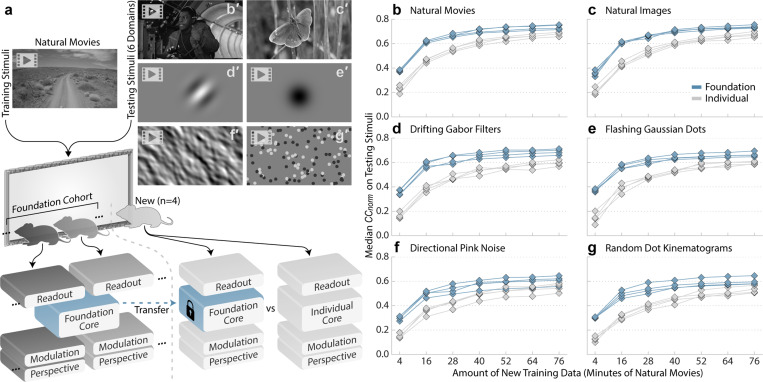
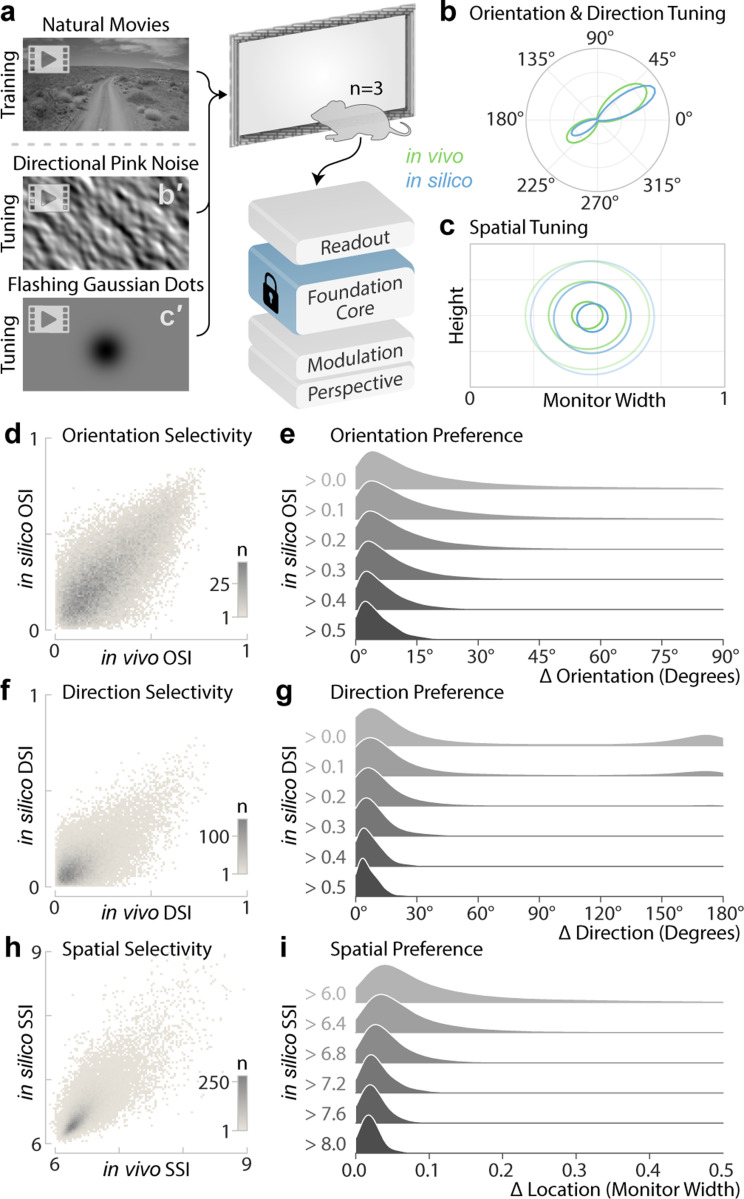
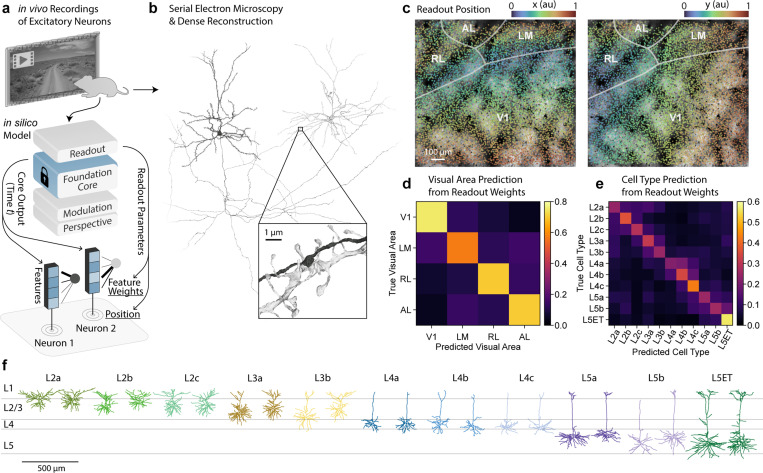
The complexity of neural circuits makes it challenging to decipher the brain's algorithms of intelligence. Recent breakthroughs in deep learning have produced models that accurately simulate brain activity, enhancing our understanding of the brain's computational objectives and neural coding. However, these models struggle to generalize beyond their training distribution, limiting their utility. The emergence of foundation models, trained on vast datasets, has introduced a new AI paradigm with remarkable generalization capabilities. We collected large amounts of neural activity from visual cortices of multiple mice and trained a foundation model to accurately predict neuronal responses to arbitrary natural videos. This model generalized to new mice with minimal training and successfully predicted responses across various new stimulus domains, such as coherent motion and noise patterns. It could also be adapted to new tasks beyond neural prediction, accurately predicting anatomical cell types, dendritic features, and neuronal connectivity within the MICrONS functional connectomics dataset. Our work is a crucial step toward building foundation brain models. As neuroscience accumulates larger, multi-modal datasets, foundation models will uncover statistical regularities, enabling rapid adaptation to new tasks and accelerating research.
Keywords: Artificial Intelligence; Foundation model; Generalization; Visual cortex.
Figures

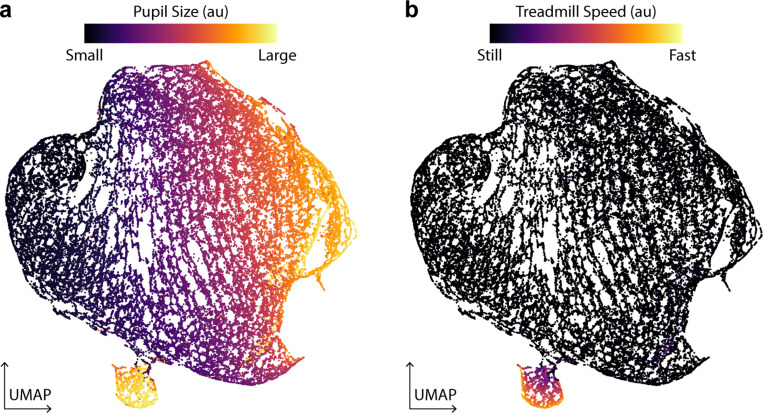



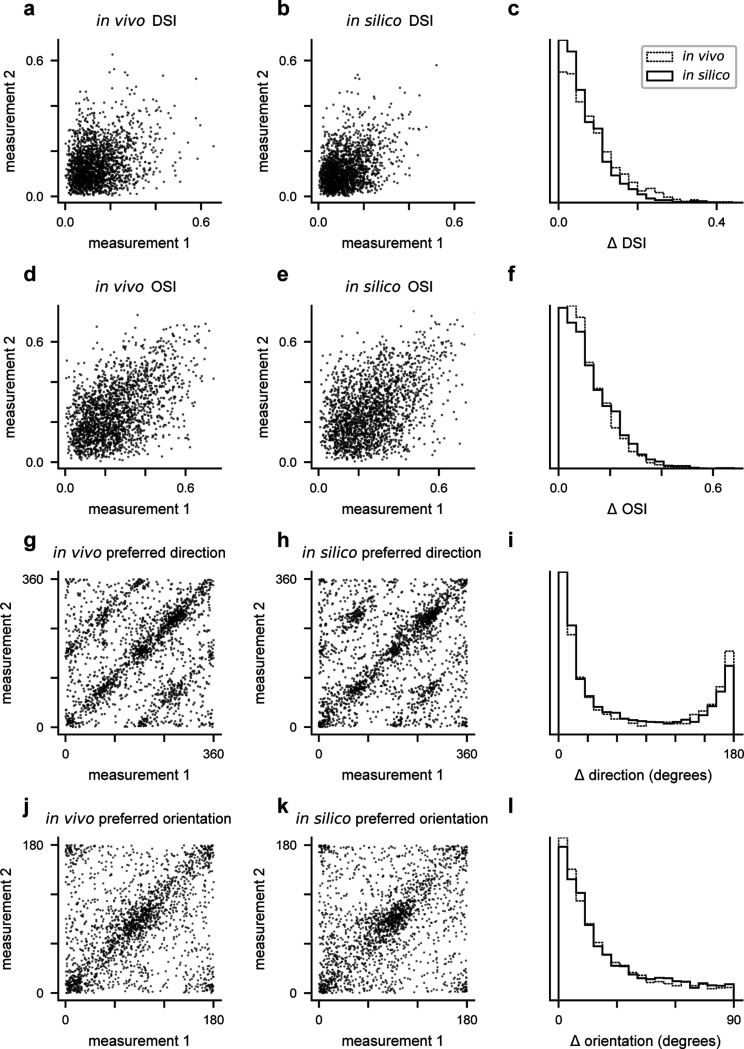

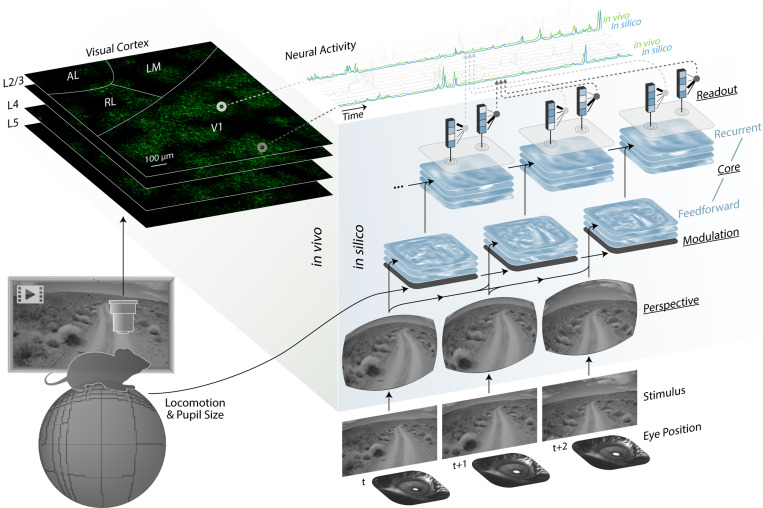
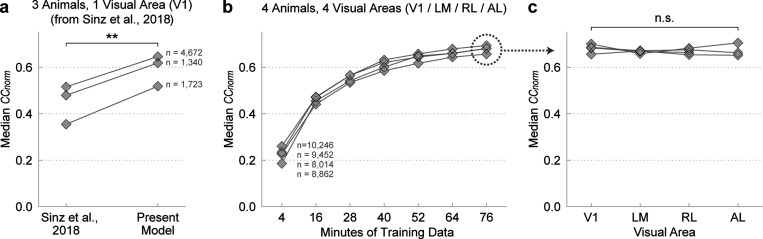
References
-
- Bakhtiari S., Mineault P, Lillicrap T., Pack C., and Richards B.. The functional specialization of visual cortex emerges from training parallel pathways with self-supervised predictive learning. Advances in Neural Information Processing Systems, 34:25164–25178,2021.
-
- Bashiri M., Walker E., Lurz K.-K., Jagadish A., Muhammad T., Ding Z., Ding Z., Tolias A., and Sinz F.. A flow-based latent state generative model of neural population responses to natural images. Advances in Neural Information Processing Systems, 34, 2021.
-
- Batty E., Merel J., Brackbill N., Heitman A., Sher A., Litke A., Chichilnisky E. J., and Paninski L.. Multilayer network models of primate retinal ganglion cells. In Proceedings of the International Conference for Learning Representations (ICLR), 2017.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources