

This is a preprint.
Genipin Crosslinks the Extracellular Matrix to Rescue Developmental and Degenerative Defects, and Accelerates Regeneration of Peripheral Neurons
- PMID: 36993570
- PMCID: PMC10055431
- DOI: 10.1101/2023.03.22.533831
Genipin Crosslinks the Extracellular Matrix to Rescue Developmental and Degenerative Defects, and Accelerates Regeneration of Peripheral Neurons
Update in
-
Genipin rescues developmental and degenerative defects in familial dysautonomia models and accelerates axon regeneration.Sci Transl Med. 2024 Nov 20;16(774):eadq2418. doi: 10.1126/scitranslmed.adq2418. Epub 2024 Nov 20. Sci Transl Med. 2024. PMID: 39565876
Abstract
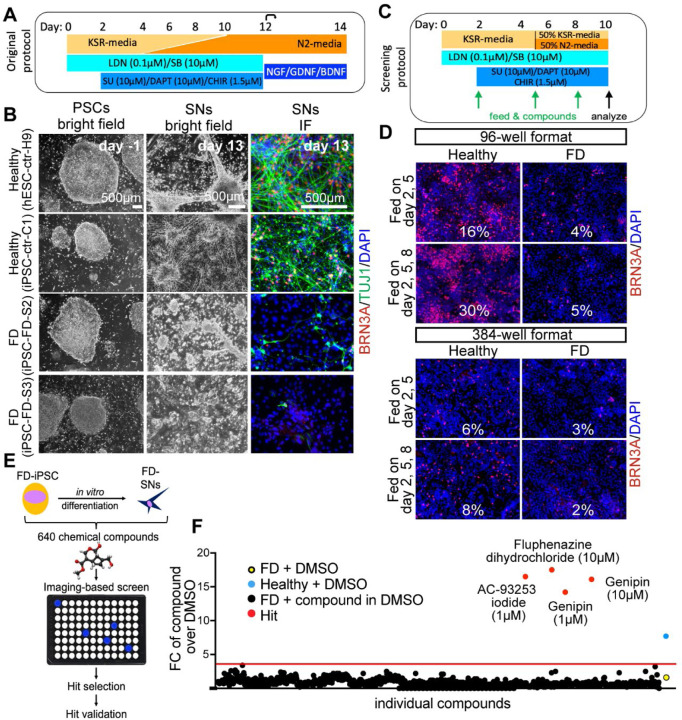
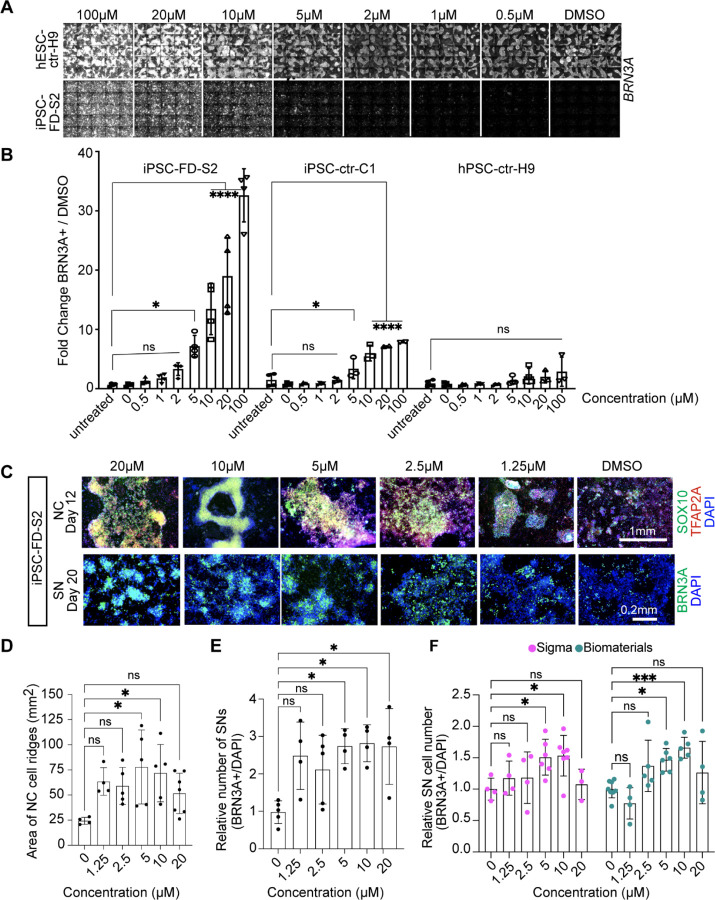
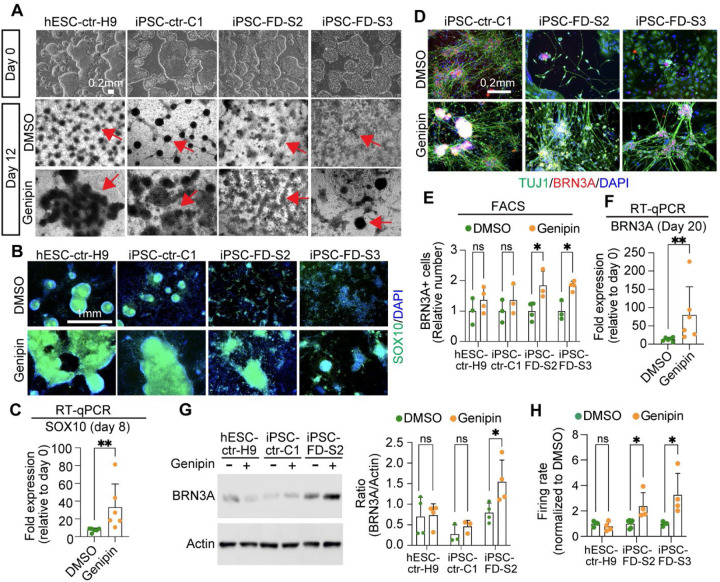
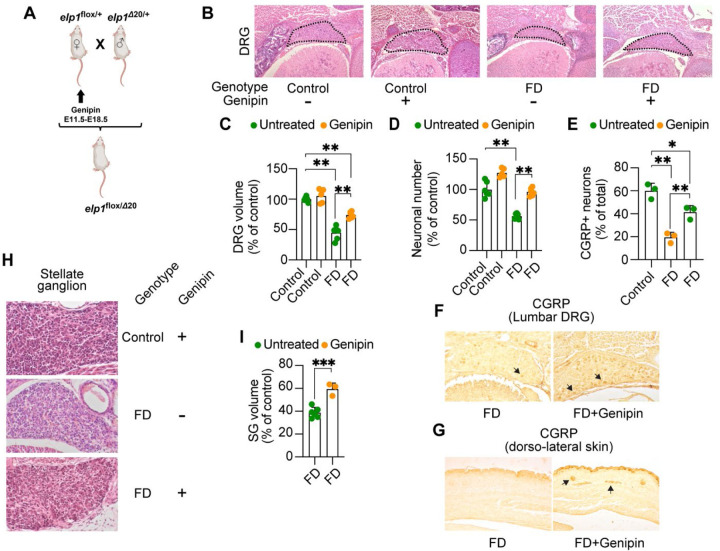
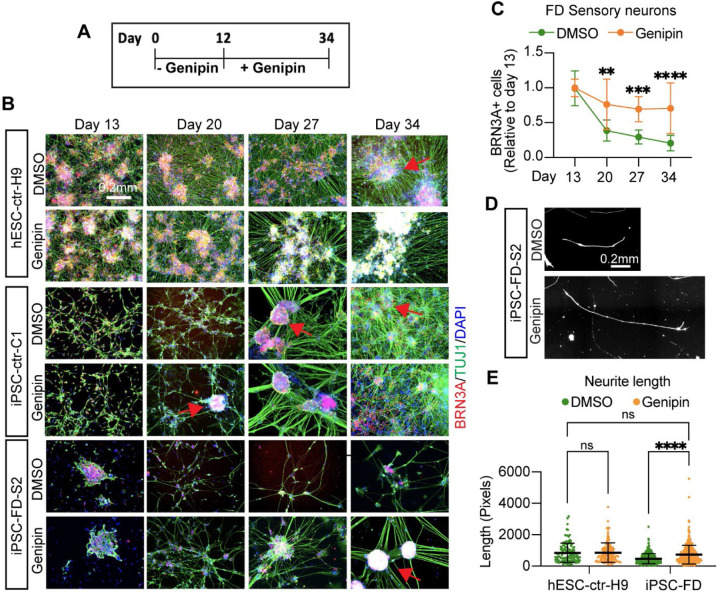
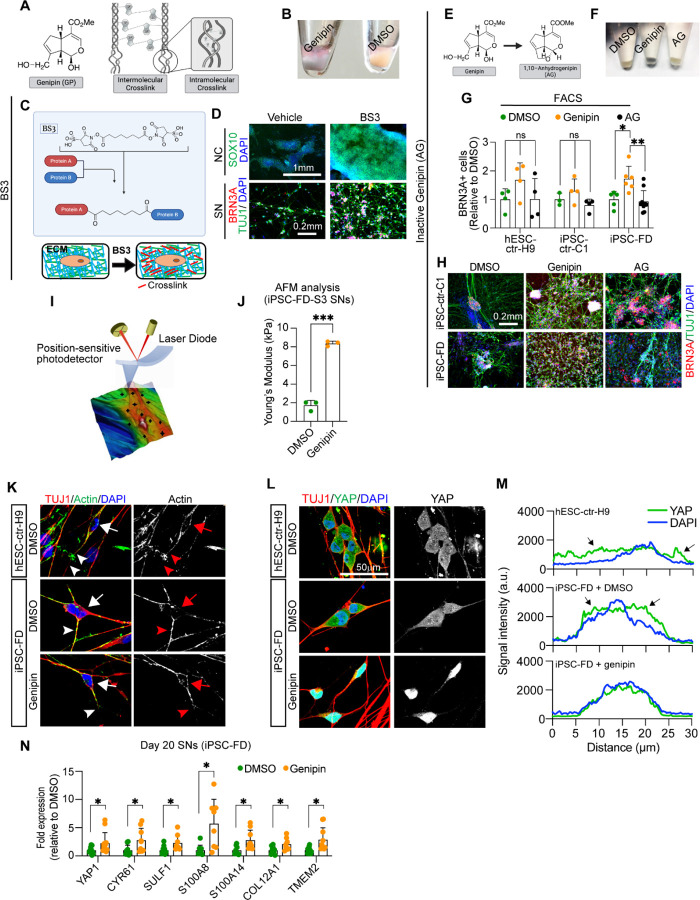
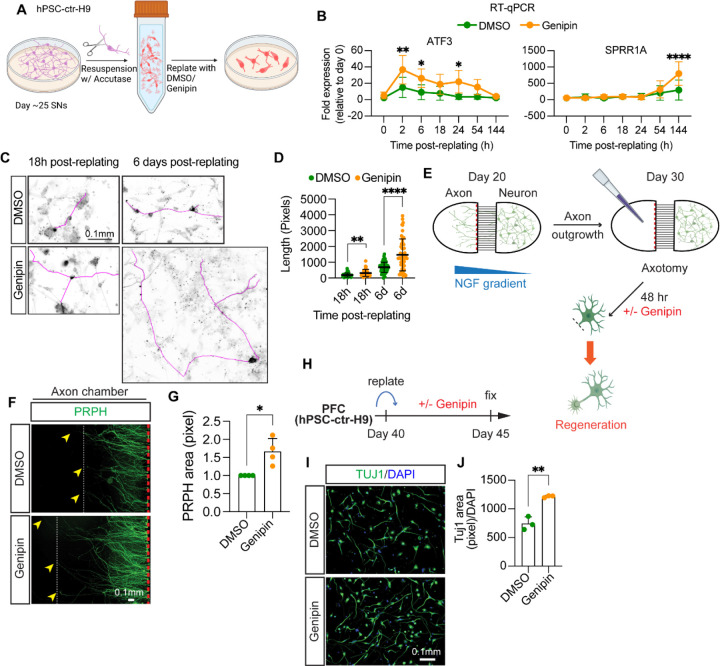
The peripheral nervous system (PNS) is essential for proper body function. A high percentage of the population suffer nerve degeneration or peripheral damage. For example, over 40% of patients with diabetes or undergoing chemotherapy develop peripheral neuropathies. Despite this, there are major gaps in the knowledge of human PNS development and therefore, there are no available treatments. Familial Dysautonomia (FD) is a devastating disorder that specifically affects the PNS making it an ideal model to study PNS dysfunction. FD is caused by a homozygous point mutation in ELP1 leading to developmental and degenerative defects in the sensory and autonomic lineages. We previously employed human pluripotent stem cells (hPSCs) to show that peripheral sensory neurons (SNs) are not generated efficiently and degenerate over time in FD. Here, we conducted a chemical screen to identify compounds able to rescue this SN differentiation inefficiency. We identified that genipin, a compound prescribed in Traditional Chinese Medicine for neurodegenerative disorders, restores neural crest and SN development in FD, both in the hPSC model and in a FD mouse model. Additionally, genipin prevented FD neuronal degeneration, suggesting that it could be offered to patients suffering from PNS neurodegenerative disorders. We found that genipin crosslinks the extracellular matrix, increases the stiffness of the ECM, reorganizes the actin cytoskeleton, and promotes transcription of YAP-dependent genes. Finally, we show that genipin enhances axon regeneration in an in vitro axotomy model in healthy sensory and sympathetic neurons (part of the PNS) and in prefrontal cortical neurons (part of the central nervous system, CNS). Our results suggest genipin can be used as a promising drug candidate for treatment of neurodevelopmental and neurodegenerative diseases, and as a enhancer of neuronal regeneration.
Conflict of interest statement
Competing interests: L.S. is a scientific co-founder and consultant and has received research support from BlueRock Therapeutics.
Figures
References
-
- Meng J., Zhang Q., Yang C., Xiao L., Xue Z., and Zhu J. (2019). Duloxetine, a Balanced Serotonin-Norepinephrine Reuptake Inhibitor, Improves Painful Chemotherapy-Induced Peripheral Neuropathy by Inhibiting Activation of p38 MAPK and NF-κB. Front Pharmacol 10, 365. 10.3389/fphar.2019.00365. - DOI - PMC - PubMed
-
- Blumenfeld A., Slaugenhaupt S.A., Liebert C.B., Temper V., Maayan C., Gill S., Lucente D.E., Idelson M., MacCormack K., Monahan M.A., et al. (1999). Precise genetic mapping and haplotype analysis of the familial dysautonomia gene on human chromosome 9q31. Am J Hum Genet 64, 1110–1118. 10.1086/302339. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources