

FN (Fibronectin)-Integrin α5 Signaling Promotes Thoracic Aortic Aneurysm in a Mouse Model of Marfan Syndrome
- PMID: 36994727
- PMCID: PMC10133209
- DOI: 10.1161/ATVBAHA.123.319120
FN (Fibronectin)-Integrin α5 Signaling Promotes Thoracic Aortic Aneurysm in a Mouse Model of Marfan Syndrome
Abstract
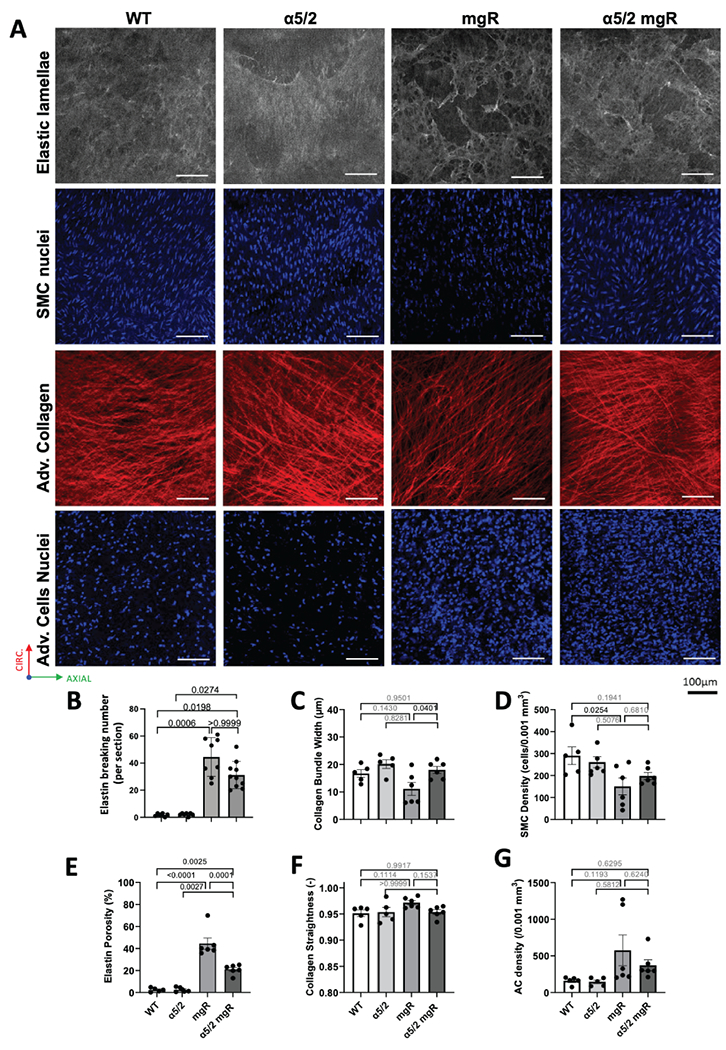
Background: Marfan syndrome, caused by mutations in the gene for fibrillin-1, leads to thoracic aortic aneurysms (TAAs). Phenotypic modulation of vascular smooth muscle cells (SMCs) and ECM (extracellular matrix) remodeling are characteristic of both nonsyndromic and Marfan aneurysms. The ECM protein FN (fibronectin) is elevated in the tunica media of TAAs and amplifies inflammatory signaling in endothelial and SMCs through its main receptor, integrin α5β1. We investigated the role of integrin α5-specific signals in Marfan mice in which the cytoplasmic domain of integrin α5 was replaced with that of integrin α2 (denoted α5/2 chimera).
Methods: We crossed α5/2 chimeric mice with Fbn1mgR/mgR mice (mgR model of Marfan syndrome) to evaluate the survival rate and pathogenesis of TAAs among wild-type, α5/2, mgR, and α5/2 mgR mice. Further biochemical and microscopic analysis of porcine and mouse aortic SMCs investigated molecular mechanisms by which FN affects SMCs and subsequent development of TAAs.
Results: FN was elevated in the thoracic aortas from Marfan patients, in nonsyndromic aneurysms, and in mgR mice. The α5/2 mutation greatly prolonged survival of Marfan mice, with improved elastic fiber integrity, mechanical properties, SMC density, and SMC contractile gene expression. Furthermore, plating of wild-type SMCs on FN decreased contractile gene expression and activated inflammatory pathways whereas α5/2 SMCs were resistant. These effects correlated with increased NF-kB activation in cultured SMCs and mgR aortas, which was alleviated by the α5/2 mutation or NF-kB inhibition.
Conclusions: FN-integrin α5 signaling is a significant driver of TAA in the mgR mouse model. This pathway thus warrants further investigation as a therapeutic target.
Keywords: fibronectin; inflammation; smooth muscle cell; target; thoracic aneurysm.
Conflict of interest statement
DISCLOSURES
JDH is a member of the Scientific Advisory Board of and paid consultant for Heartflow, Inc., an AI-based company focused on non-invasive assessment of coronary artery disease. The other authors declare no conflicts, financial or otherwise.
Figures






References
-
- Sengle G and Sakai LY. The fibrillin microfibril scaffold: A niche for growth factors and mechanosensation? Matrix Biol. 2015;47:3–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
