

Base editing rescue of spinal muscular atrophy in cells and in mice
- PMID: 36996170
- PMCID: PMC10270003
- DOI: 10.1126/science.adg6518
Base editing rescue of spinal muscular atrophy in cells and in mice
Abstract
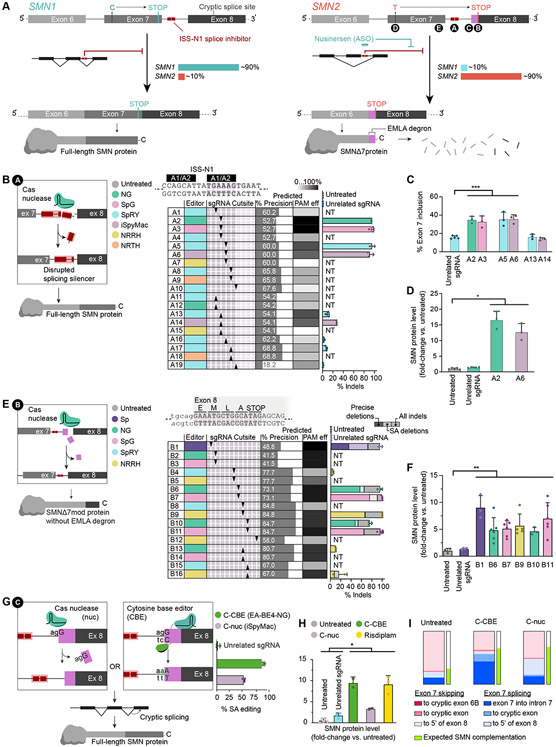
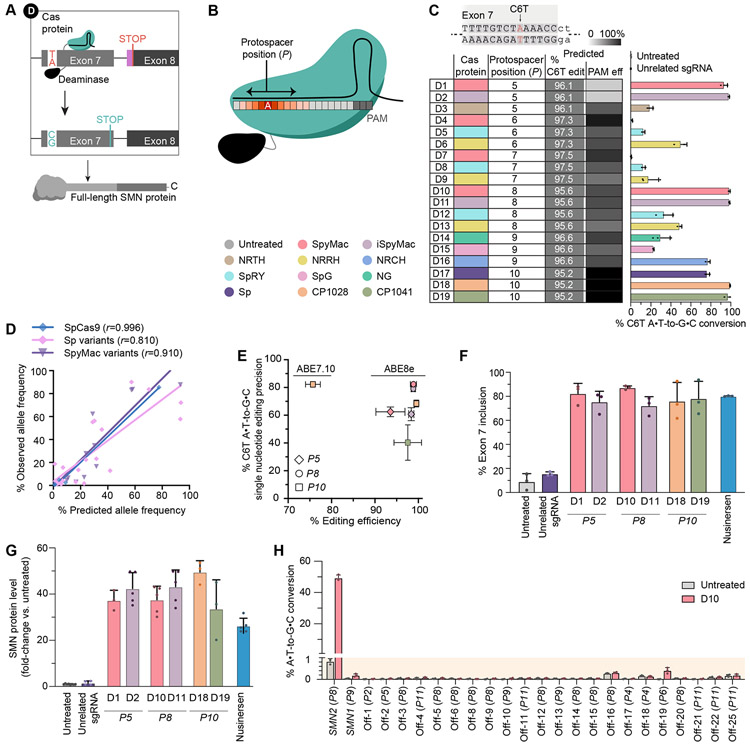
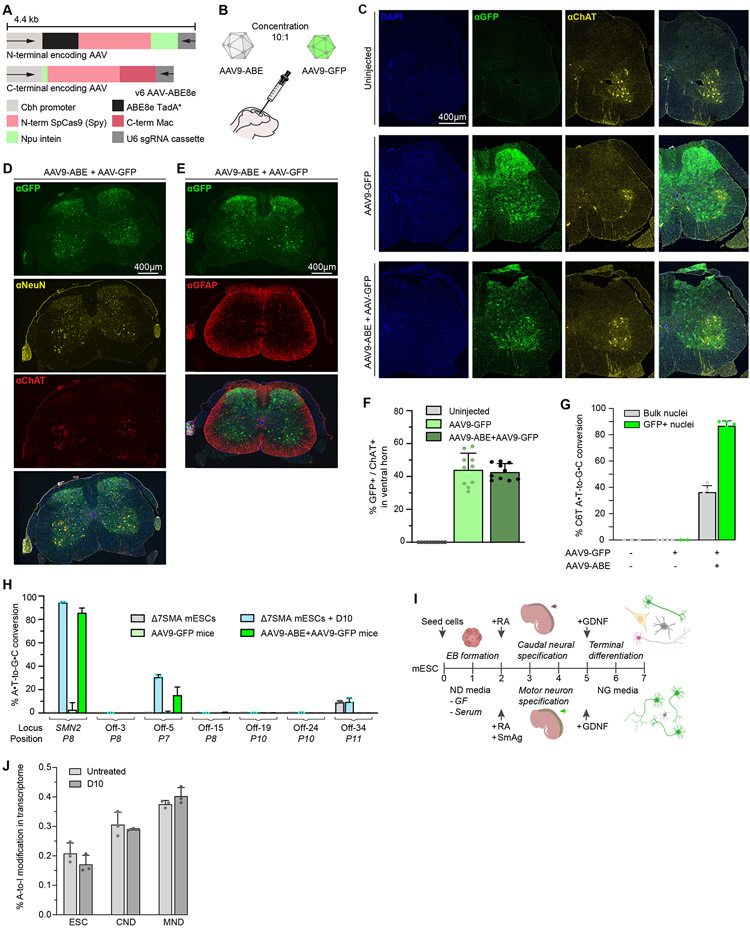
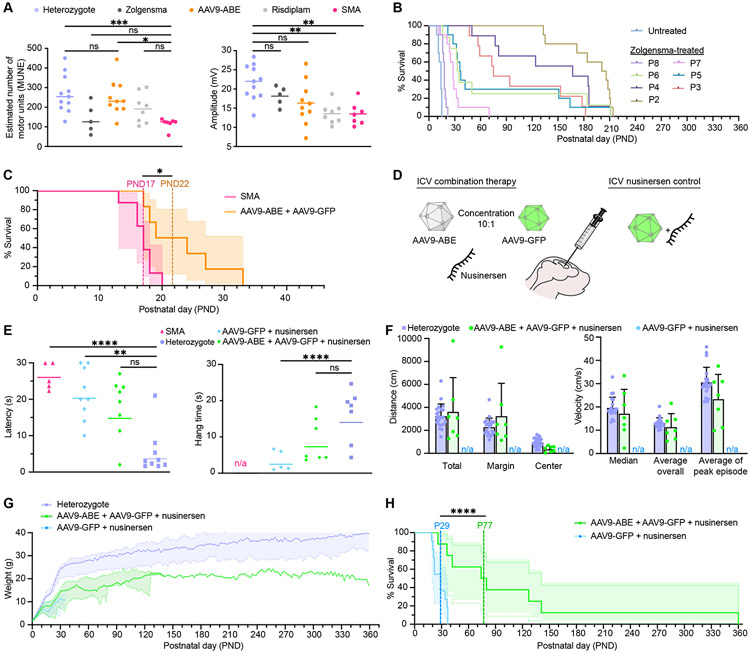
Spinal muscular atrophy (SMA), the leading genetic cause of infant mortality, arises from survival motor neuron (SMN) protein insufficiency resulting from SMN1 loss. Approved therapies circumvent endogenous SMN regulation and require repeated dosing or may wane. We describe genome editing of SMN2, an insufficient copy of SMN1 harboring a C6>T mutation, to permanently restore SMN protein levels and rescue SMA phenotypes. We used nucleases or base editors to modify five SMN2 regulatory regions. Base editing converted SMN2 T6>C, restoring SMN protein levels to wild type. Adeno-associated virus serotype 9-mediated base editor delivery in Δ7SMA mice yielded 87% average T6>C conversion, improved motor function, and extended average life span, which was enhanced by one-time base editor and nusinersen coadministration (111 versus 17 days untreated). These findings demonstrate the potential of a one-time base editing treatment for SMA.
Figures
Comment in
-
Base editing treats SMA in mice.Nat Rev Drug Discov. 2023 May;22(5):353. doi: 10.1038/d41573-023-00057-1. Nat Rev Drug Discov. 2023. PMID: 37024675 No abstract available.
References
-
- Lefebvre S, Bürglen L, Reboullet S, Clermont O, Burlet P, Viollet L, Benichou B, Cruaud C, Millasseau P, Zeviani M, Identification and characterization of a spinal muscular atrophy-determining gene. Cell. 80, 155–65 (1995). - PubMed
-
- Sugarman EA, Nagan N, Zhu H, Akmaev VR, Zhou Z, Rohlfs EM, Flynn K, Hendrickson BC, Scholl T, Sirko-Osadsa DA, Allitto BA, Pan-ethnic carrier screening and prenatal diagnosis for spinal muscular atrophy: clinical laboratory analysis of > 72 400 specimens. Eur. J. Hum. Genet 20, 27–32 (2012). - PMC - PubMed
-
- Boda B, Mas C, Giudicelli C, Nepote V, Guimiot F, Levacher B, Zvara A, Santha M, LeGall I, Simonneau M, Survival motor neuron SMN1 and SMN2 gene promoters: Identical sequences and differential expression in neurons and non-neuronal cells. Eur. J. Hum. Genet 12, 729–737 (2004). - PubMed
-
- Rochette CF, Gilbert N, Simard LR, SMN gene duplication and the emergence of the SMN2 gene occurred in distinct hominids: SMN2 is unique to Homo sapiens. Hum. Genet 108, 255–266 (2001). - PubMed
MeSH terms
Substances
Grants and funding
- R00 NS119743/NS/NINDS NIH HHS/United States
- K99 HL163805/HL/NHLBI NIH HHS/United States
- R00 HL163805/HL/NHLBI NIH HHS/United States
- R35 GM118062/GM/NIGMS NIH HHS/United States
- R01 HD060586/HD/NICHD NIH HHS/United States
- R01 EB022376/EB/NIBIB NIH HHS/United States
- P01 HL053749/HL/NHLBI NIH HHS/United States
- RM1 HG009490/HG/NHGRI NIH HHS/United States
- K99 NS119743/NS/NINDS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- T32 GM095450/GM/NIGMS NIH HHS/United States
- U01 AI142756/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
