

Protein disulfide-isomerase A4 confers glioblastoma angiogenesis promotion capacity and resistance to anti-angiogenic therapy
- PMID: 36997943
- PMCID: PMC10061982
- DOI: 10.1186/s13046-023-02640-1
Protein disulfide-isomerase A4 confers glioblastoma angiogenesis promotion capacity and resistance to anti-angiogenic therapy
Abstract
Introduction: Increasing evidence has revealed the key activity of protein disulfide isomerase A4 (PDIA4) in the endoplasmic reticulum stress (ERS) response. However, the role of PDIA4 in regulating glioblastoma (GBM)-specific pro-angiogenesis is still unknown.
Methods: The expression and prognostic role of PDIA4 were analyzed using a bioinformatics approach and were validated in 32 clinical samples and follow-up data. RNA-sequencing was used to search for PDIA4-associated biological processes in GBM cells, and proteomic mass spectrum (MS) analysis was used to screen for potential PDIA4 substrates. Western blotting, real-time quantitative polymerase chain reaction (RT-qPCR), and enzyme-linked immunosorbent assays (ELISA) were used to measure the levels of the involved factors. Cell migration and tube formation assays determined the pro-angiogenesis activity of PDIA4 in vitro. An intracranial U87 xenograft GBM animal model was constructed to evaluate the pro-angiogenesis role of PDIA4 in vivo.
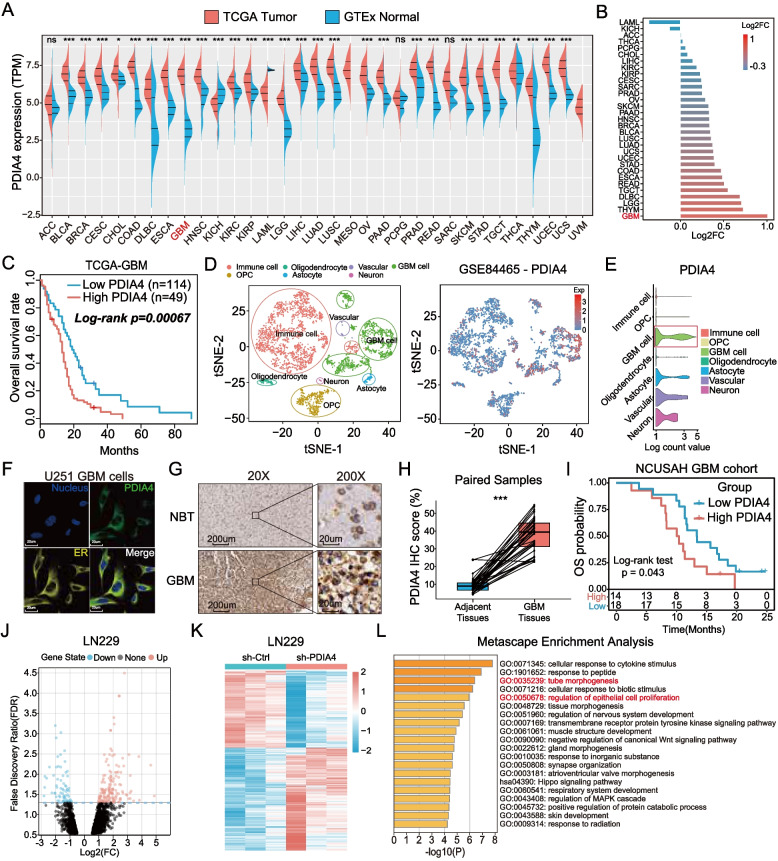
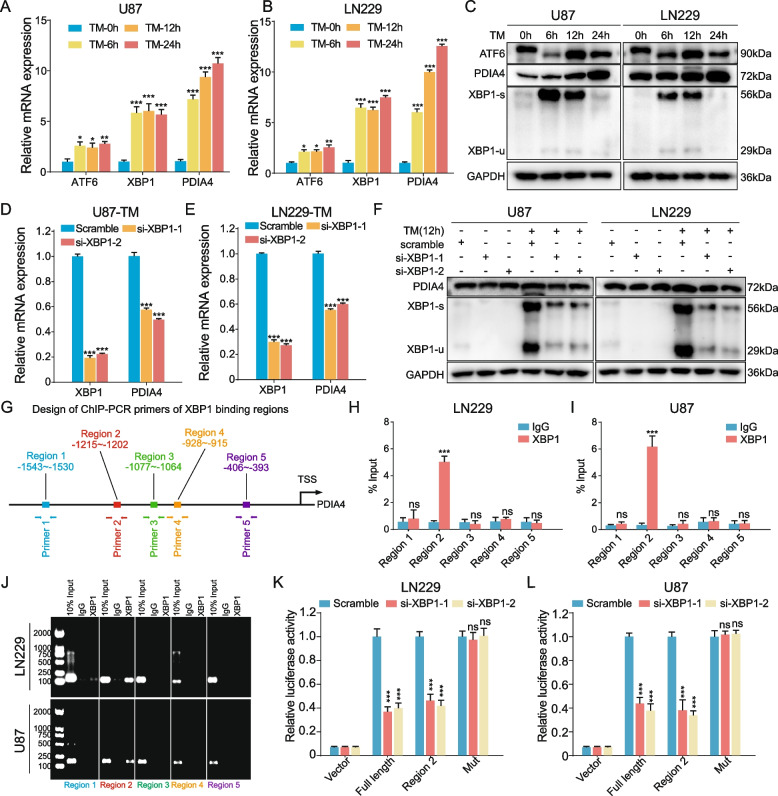
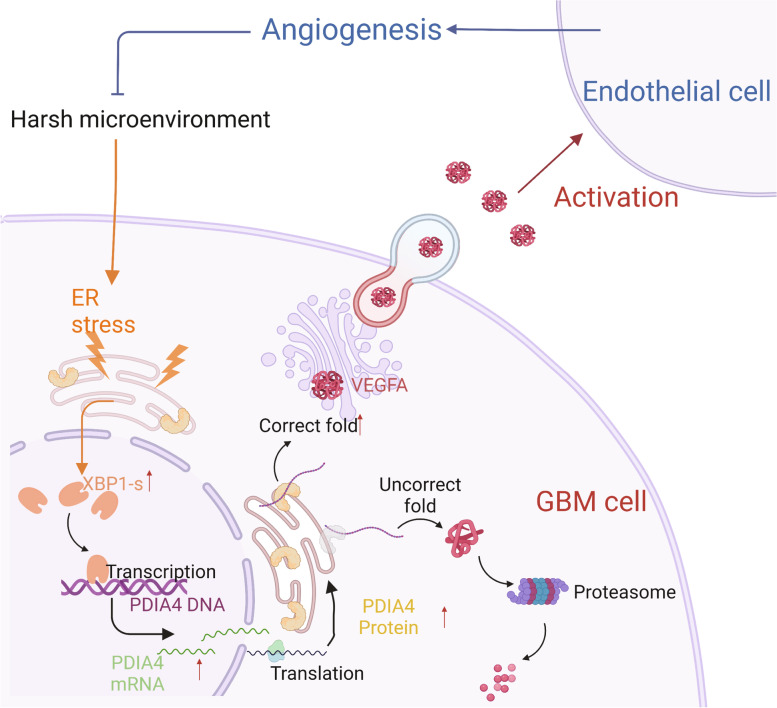
Results: Aberrant overexpression of PDIA4 was associated with a poor prognosis in patients with GBM, although PDIA4 could also functionally regulate intrinsic GBM secretion of vascular endothelial growth factor-A (VEGF-A) through its active domains of Cys-X-X-Cys (CXXC) oxidoreductase. Functionally, PDIA4 exhibits pro-angiogenesis activity both in vitro and in vivo, and can be upregulated by ERS through transcriptional regulation of X-box binding protein 1 (XBP1). The XBP1/PDIA4/VEGFA axis partially supports the mechanism underlying GBM cell survival under ER stress. Further, GBM cells with higher expression of PDIA4 showed resistance to antiangiogenic therapy in vivo.
Conclusions: Our findings revealed the pro-angiogenesis role of PDIA4 in GBM progression and its potential impact on GBM survival under a harsh microenvironment. Targeting PDIA4 might help to improve the efficacy of antiangiogenic therapy in patients with GBM.
Keywords: Angiogenesis; Endoplasmic reticulum stress (ERS); Glioblastoma (GBM); Protein disulfide-isomerase A4 (PDIA4); X-box binding protein 1 (XBP1).
© 2023. The Author(s).
Conflict of interest statement
No competing interest exists.
Figures




References
MeSH terms
Substances
Grants and funding
- 81960456/National Natural Science Foundation of China
- 82172989/National Natural Science Foundation of China
- 82273068/National Natural Science Foundation of China
- 82260524/National Natural Science Foundation of China
- 20203CCH45008/Introduced and jointly Built High-end R&D Institute of Jiangxi
- 20212BBG73021/Key Research and Development projects in Jiangxi
- 2023ZD003/Key project of Science and Technology Innovation of Health Commission
- GJJ210177/Jiangxi Province Department of Education Science and technology research project, China
- 20212BCJ23023/Jiangxi Training Program for academic and technical leaders of major disciplines -- Young talents program
- 20192BAB205077/Natural Science Foundation of Jiangxi Province
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
