

Methylglyoxal: a novel upstream regulator of DNA methylation
- PMID: 36998085
- PMCID: PMC10064647
- DOI: 10.1186/s13046-023-02637-w
Methylglyoxal: a novel upstream regulator of DNA methylation
Abstract
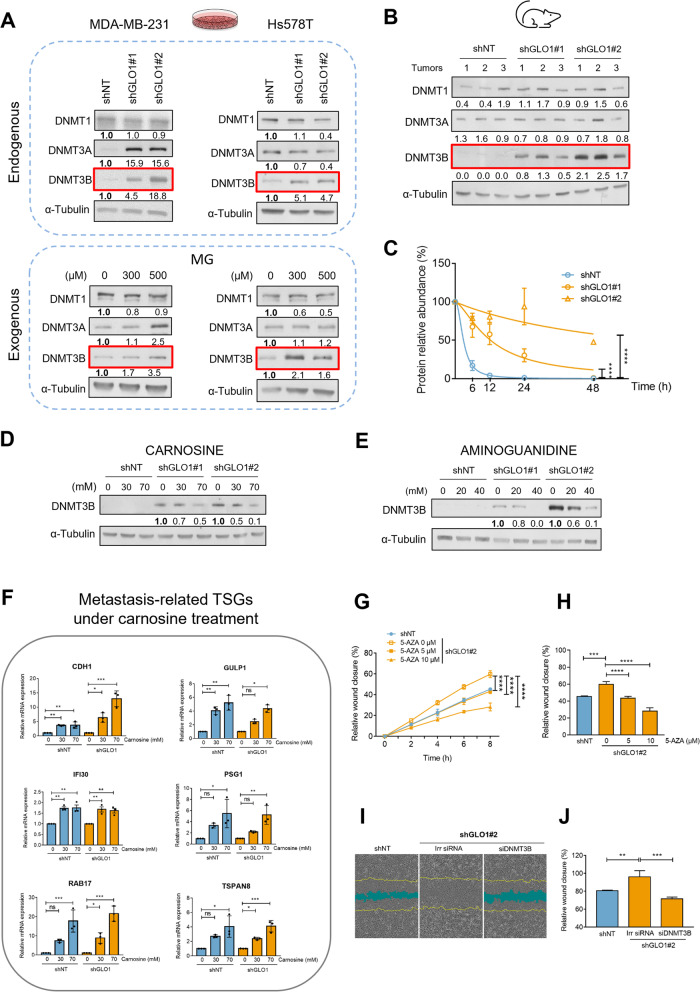
Background: Aerobic glycolysis, also known as the Warburg effect, is predominantly upregulated in a variety of solid tumors, including breast cancer. We have previously reported that methylglyoxal (MG), a very reactive by-product of glycolysis, unexpectedly enhanced the metastatic potential in triple negative breast cancer (TNBC) cells. MG and MG-derived glycation products have been associated with various diseases, such as diabetes, neurodegenerative disorders, and cancer. Glyoxalase 1 (GLO1) exerts an anti-glycation defense by detoxifying MG to D-lactate.
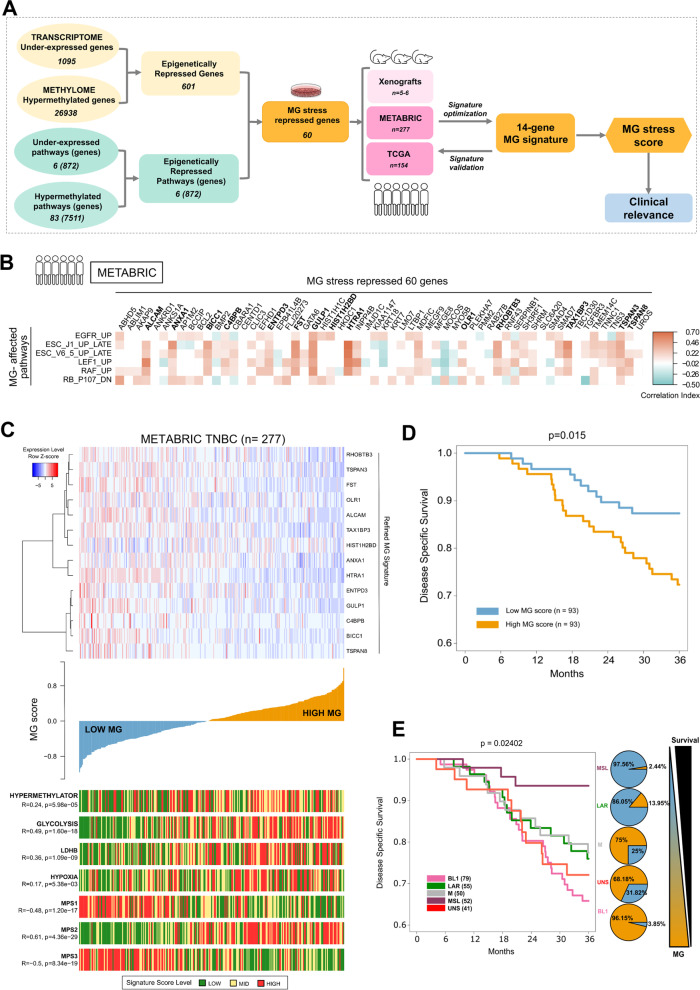
Methods: Here, we used our validated model consisting of stable GLO1 depletion to induce MG stress in TNBC cells. Using genome-scale DNA methylation analysis, we report that this condition resulted in DNA hypermethylation in TNBC cells and xenografts.
Results: GLO1-depleted breast cancer cells showed elevated expression of DNMT3B methyltransferase and significant loss of metastasis-related tumor suppressor genes, as assessed using integrated analysis of methylome and transcriptome data. Interestingly, MG scavengers revealed to be as potent as typical DNA demethylating agents at triggering the re-expression of representative silenced genes. Importantly, we delineated an epigenomic MG signature that effectively stratified TNBC patients based on survival.
Conclusion: This study emphasizes the importance of MG oncometabolite, occurring downstream of the Warburg effect, as a novel epigenetic regulator and proposes MG scavengers to reverse altered patterns of gene expression in TNBC.
Keywords: Breast cancer; DNA methylation; Metastasis; Methylglyoxal; Tumor suppressor genes.
© 2023. The Author(s).
Conflict of interest statement
None of the authors have a conflict of interest.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
