

Hypothalamic neuroglial plasticity is regulated by anti-Müllerian hormone and disrupted in polycystic ovary syndrome
- PMID: 37001236
- PMCID: PMC10070524
- DOI: 10.1016/j.ebiom.2023.104535
Hypothalamic neuroglial plasticity is regulated by anti-Müllerian hormone and disrupted in polycystic ovary syndrome
Abstract
Background: Polycystic ovary syndrome (PCOS) is the most common reproductive-endocrine disorder affecting between 5 and 18% of women worldwide. An elevated frequency of pulsatile luteinizing hormone (LH) secretion and higher serum levels of anti-Müllerian hormone (AMH) are frequently observed in women with PCOS. The origin of these abnormalities is, however, not well understood.
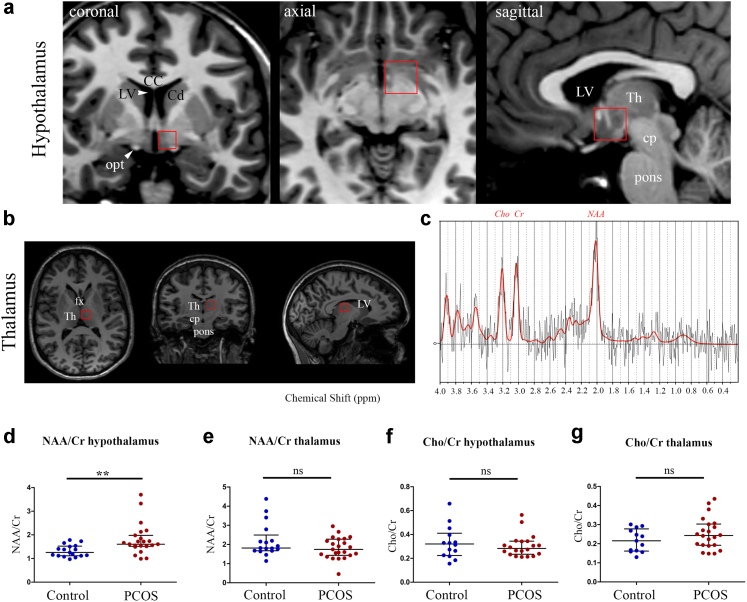
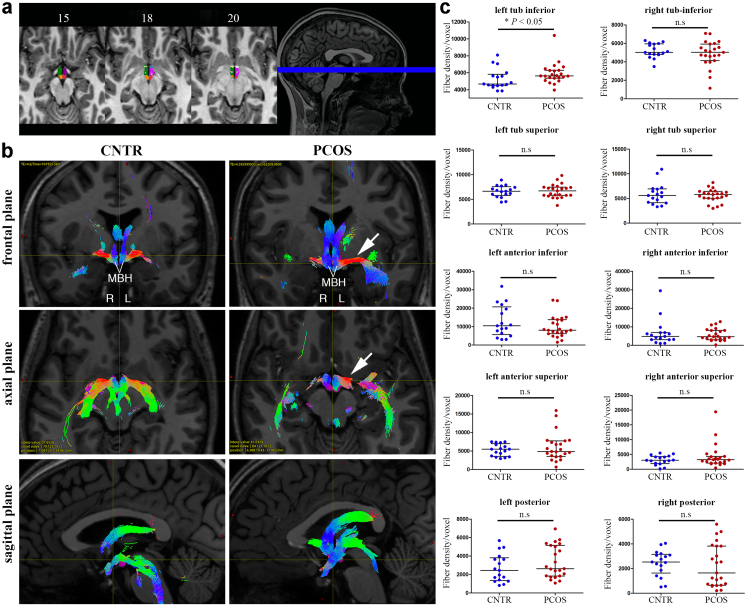
Methods: We studied brain structure and function in women with and without PCOS using proton magnetic resonance spectroscopy (MRS) and diffusion tensor imaging combined with fiber tractography. Then, using a mouse model of PCOS, we investigated by electron microscopy whether AMH played a role on the regulation of hypothalamic structural plasticity.
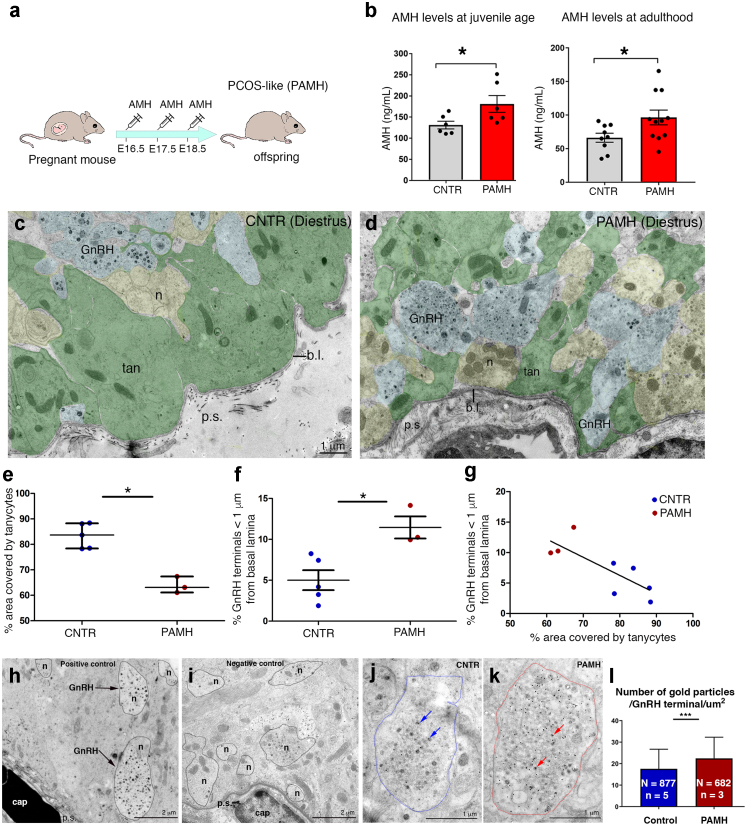
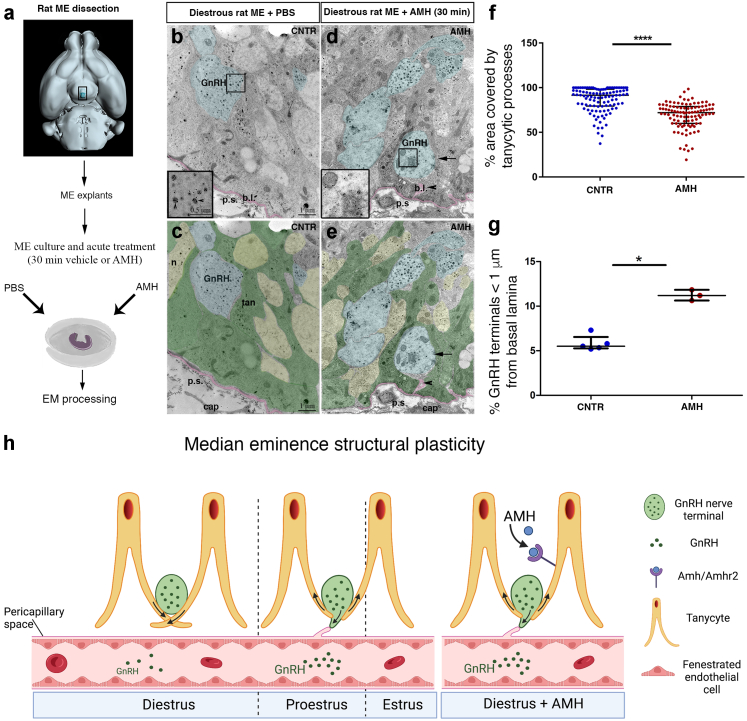
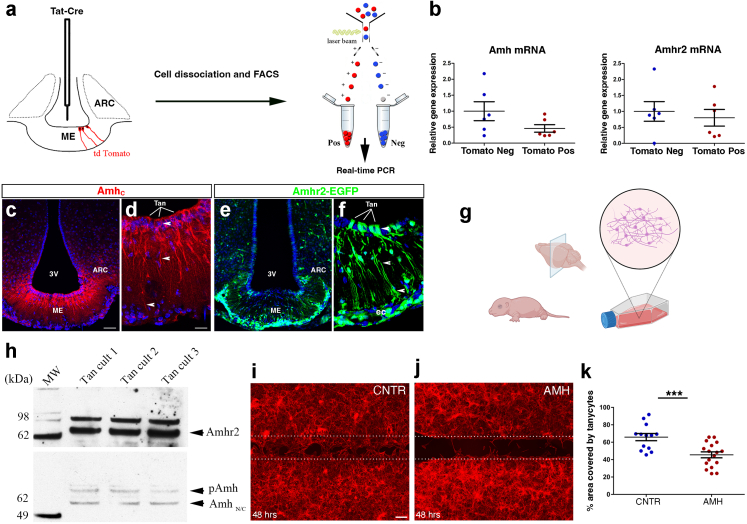
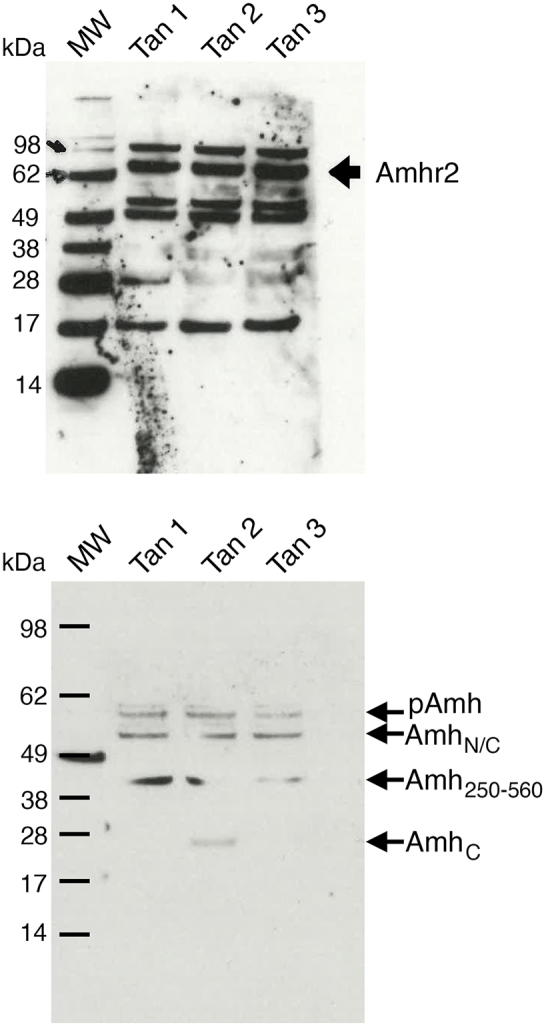
Findings: Increased AMH serum levels are associated with increased hypothalamic activity/axonal-glial signalling in PCOS patients. Furthermore, we demonstrate that AMH promotes profound micro-structural changes in the murine hypothalamic median eminence (ME), creating a permissive environment for GnRH secretion. These include the retraction of the processes of specialized AMH-sensitive ependymo-glial cells called tanycytes, allowing more GnRH neuron terminals to approach ME blood capillaries both during the run-up to ovulation and in a mouse model of PCOS.
Interpretation: We uncovered a central function for AMH in the regulation of fertility by remodeling GnRH terminals and their tanycytic sheaths, and provided insights into the pivotal role of the brain in the establishment and maintenance of neuroendocrine dysfunction in PCOS.
Funding: INSERM (U1172), European Research Council (ERC) under the European Union's Horizon 2020 research and innovation program (grant agreement n° 725149), CHU de Lille, France (Bonus H).
Keywords: AMH; GnRH; Hypothalamus; MR spectroscopy; PCOS; Tanycytes.
Copyright © 2023 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests A.-L.B. received a honorarium from Merck-Serono for a booklet writing on the use of AMH in Assisted Reproductive Technology; G.K. was a recipient of a Research Grant from Société Française de Neuroradiologie that covered part of his salary. All the other authors have declared that no conflict of interest exists.
Figures
Comment in
-
Polycystic ovary syndrome: deciphering mechanisms to facilitate management and treatment.EBioMedicine. 2023 Aug;94:104754. doi: 10.1016/j.ebiom.2023.104754. EBioMedicine. 2023. PMID: 37567728 Free PMC article. No abstract available.
References
-
- Belchetz P.E., Plant T.M., Nakai Y., Keogh E.J., Knobil E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 1978;202(4368):631–633. - PubMed
-
- Prevot V., Bellefontaine N., Baroncini M., et al. Gonadotrophin-releasing hormone nerve terminals, tanycytes and neurohaemal junction remodelling in the adult median eminence: functional consequences for reproduction and dynamic role of vascular endothelial cells. J Neuroendocrinol. 2010;22(7):639–649. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
