

MAIT cell inhibition promotes liver fibrosis regression via macrophage phenotype reprogramming
- PMID: 37005415
- PMCID: PMC10067815
- DOI: 10.1038/s41467-023-37453-5
MAIT cell inhibition promotes liver fibrosis regression via macrophage phenotype reprogramming
Abstract
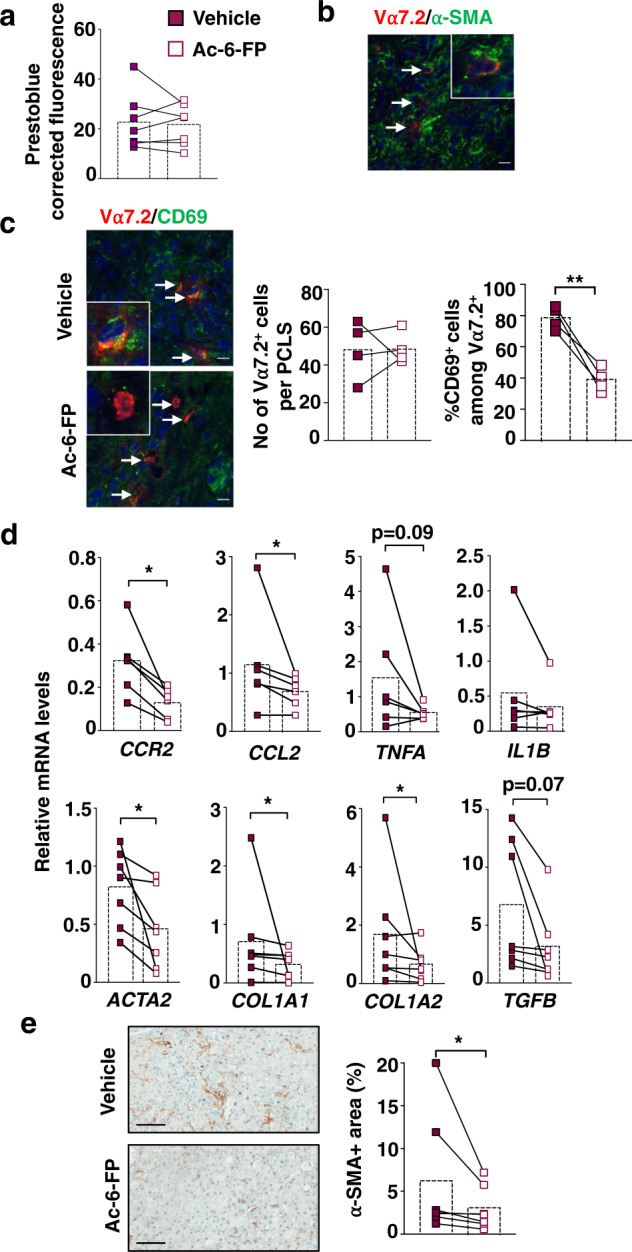
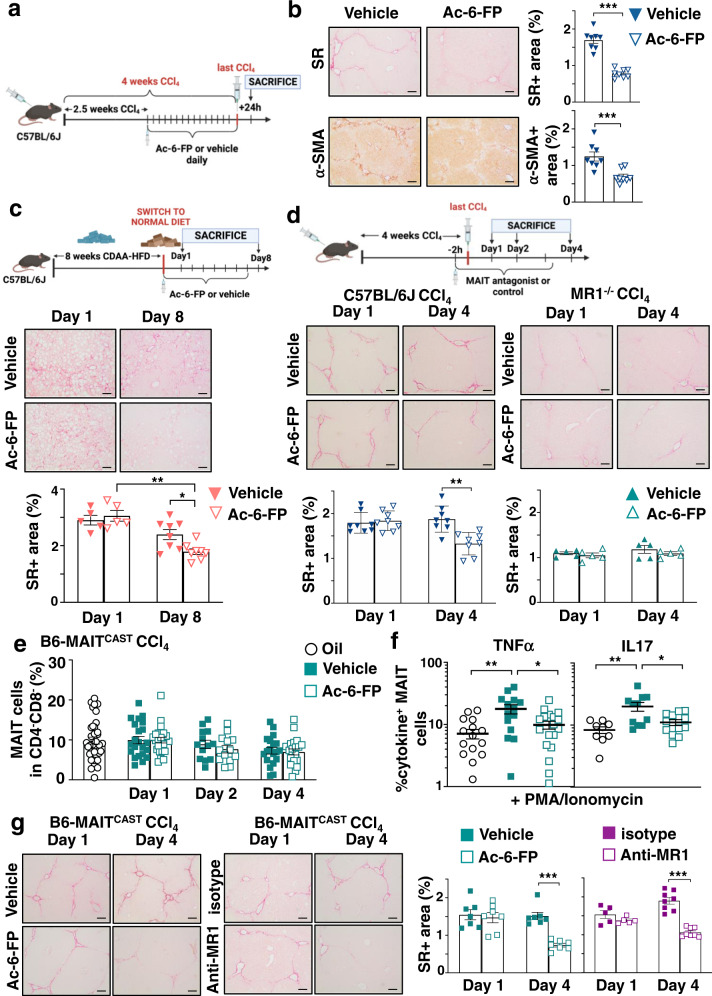
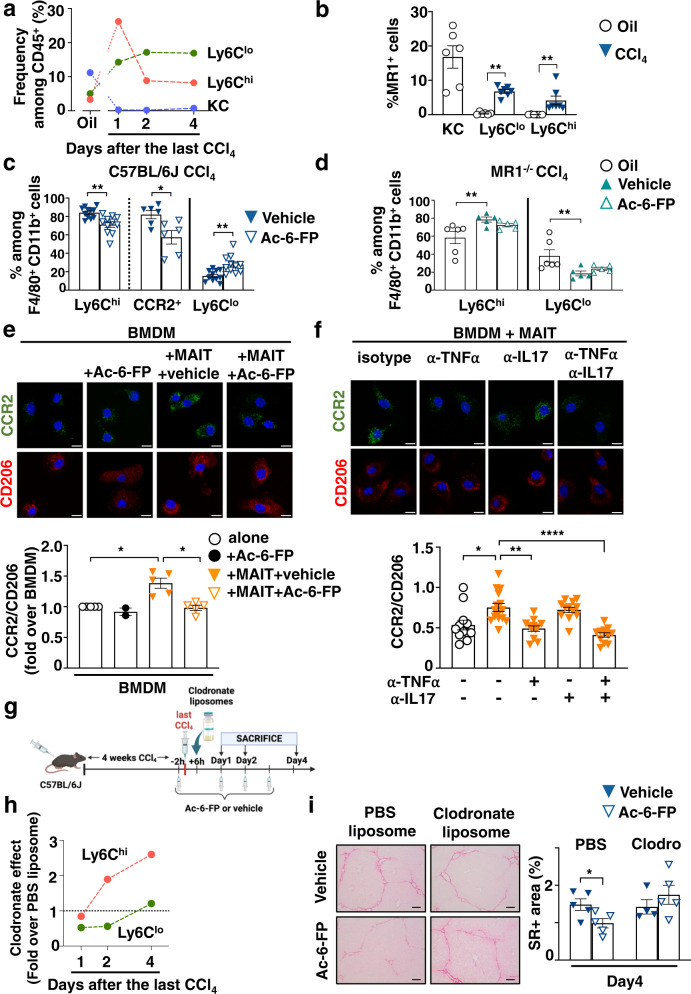
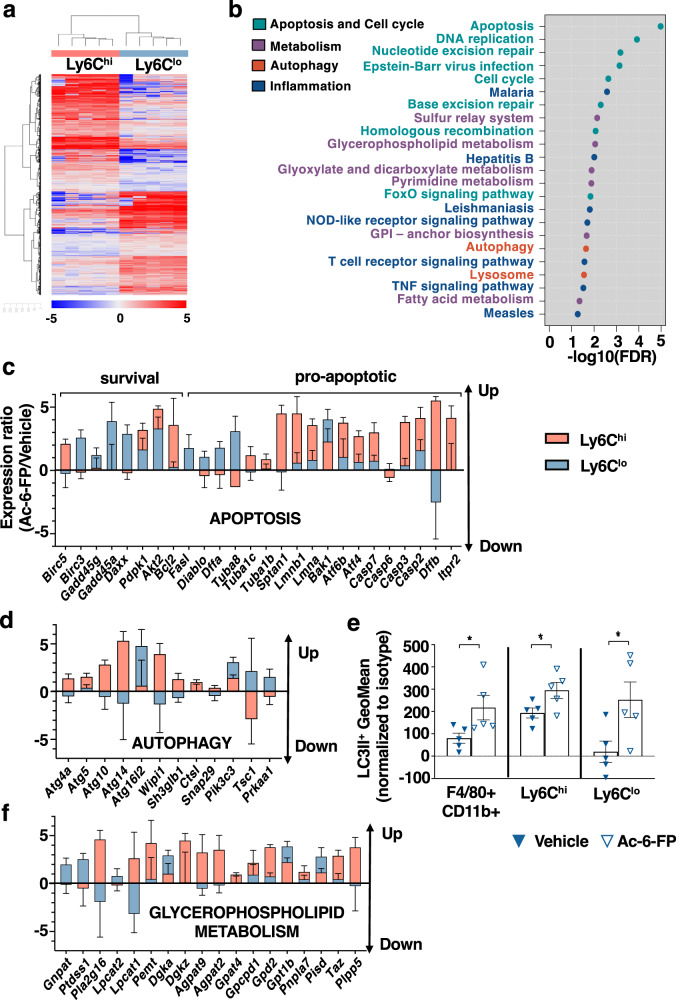
Recent data have shown that liver fibrosis can regress even at later stages of cirrhosis and shifting the immune response from pro-inflammatory towards a resolutive profile is considered as a promising option. The immune regulatory networks that govern the shift of the inflammatory phenotype and thus potential reversal of liver fibrosis are lesser known. Here we show that in precision-cut human liver slices obtained from patients with end-stage fibrosis and in mouse models, inhibiting Mucosal-Associated Invariant T (MAIT) cells using pharmacological or antibody-driven approaches, limits fibrosis progression and even regresses fibrosis, following chronic toxic- or non-alcoholic steatohepatitis (NASH)-induced liver injury. Mechanistic studies, combining RNA sequencing, in vivo functional studies (performed in male mice) and co-culture experiments indicate that disruption of the MAIT cell-monocyte/macrophage interaction results in resolution of fibrosis both by increasing the frequency of restorative Ly6Clo at the expenses of pro-fibrogenic Ly6Chi monocyte-derived macrophages and promoting an autophagic phenotype in both subsets. Thus, our data show that MAIT cell activation and the consequential phenotype shift of liver macrophages are important pathogenic features of liver fibrosis and could be targeted by anti-fibrogenic therapy.
© 2023. The Author(s).
Conflict of interest statement
All authors declare no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
