

Wnt/β-catenin signaling induces axial elasticity patterns of Hydra extracellular matrix
- PMID: 37009232
- PMCID: PMC10050647
- DOI: 10.1016/j.isci.2023.106416
Wnt/β-catenin signaling induces axial elasticity patterns of Hydra extracellular matrix
Abstract
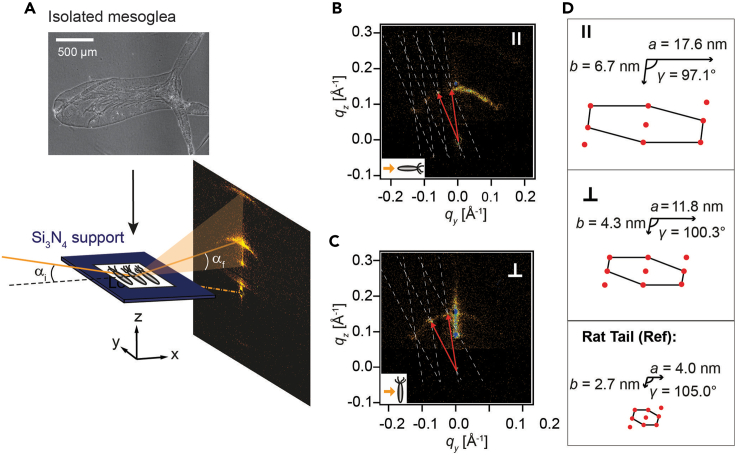
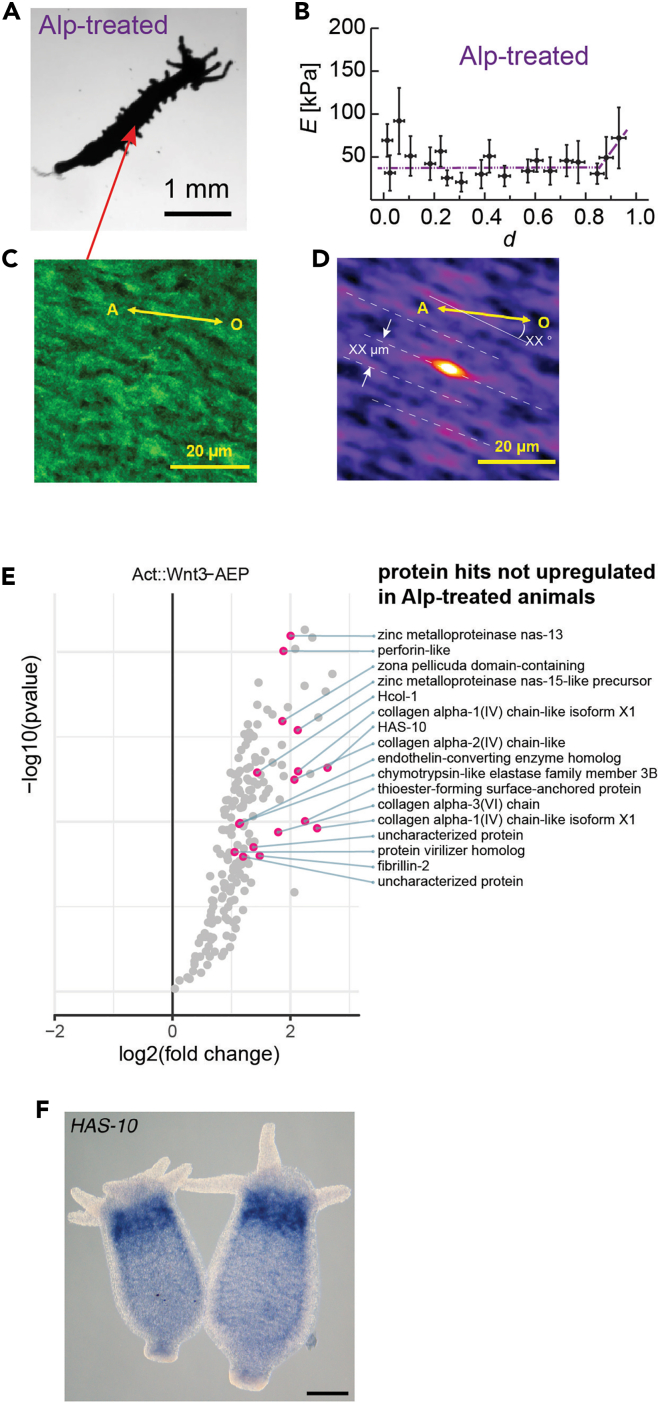
The extracellular matrix (ECM) plays crucial roles in animal development and diseases. Here, we report that Wnt/β-catenin signaling induces the ECM remodeling during Hydra axis formation. We determined the micro- and nanoscopic arrangement of fibrillar type I collagen along Hydra's body axis using high-resolution microscopy and X-ray scattering. Elasticity mapping of the ECM ex vivo revealed distinctive elasticity patterns along the body axis. A proteomic analysis of the ECM showed that these elasticity patterns correlate with a gradient-like distribution of metalloproteases along the body axis. Activation of the Wnt/β-catenin pathway in wild-type and transgenic animals alters these patterns toward low ECM elasticity patterns. This suggests a mechanism whereby high protease activity under control of Wnt/β-catenin signaling causes remodeling and softening of the ECM. This Wnt-dependent spatiotemporal coordination of biochemical and biomechanical cues in ECM formation was likely a central evolutionary innovation for animal tissue morphogenesis.
Keywords: Histology; Microscopic anatomy; Molecular biology; Zoology.
© 2023 The Authors.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
