

Viruses of a key coral symbiont exhibit temperature-driven productivity across a reefscape
- PMID: 37009785
- PMCID: PMC10068613
- DOI: 10.1038/s43705-023-00227-7
Viruses of a key coral symbiont exhibit temperature-driven productivity across a reefscape
Abstract
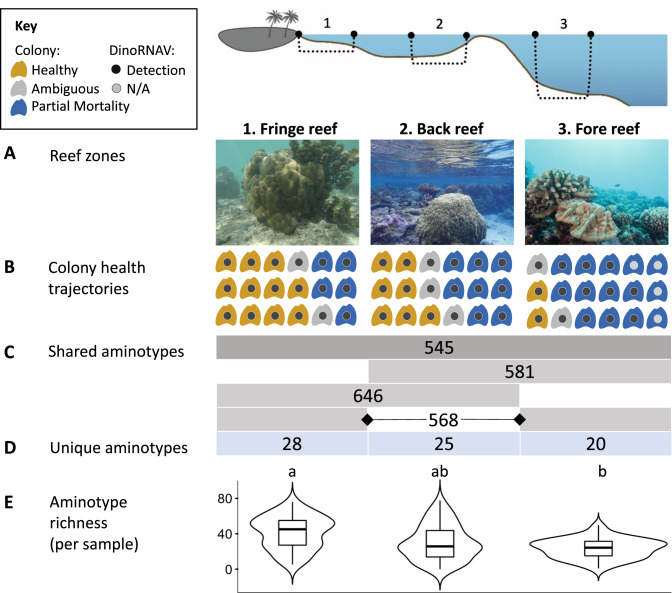
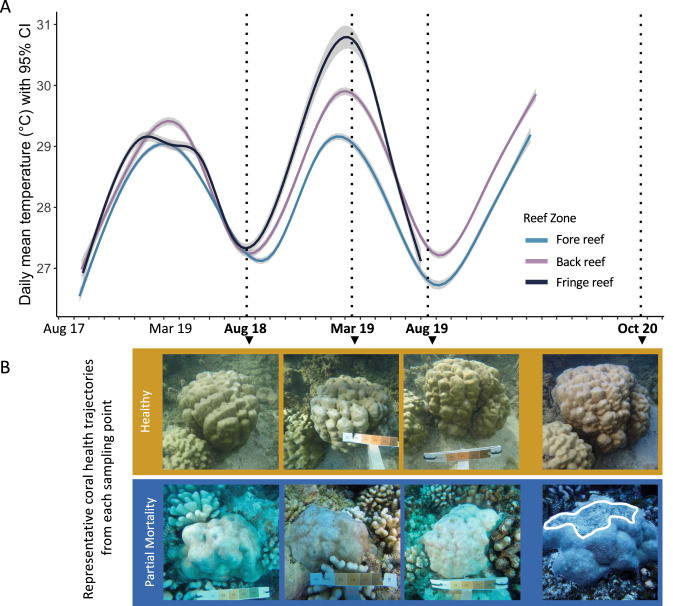
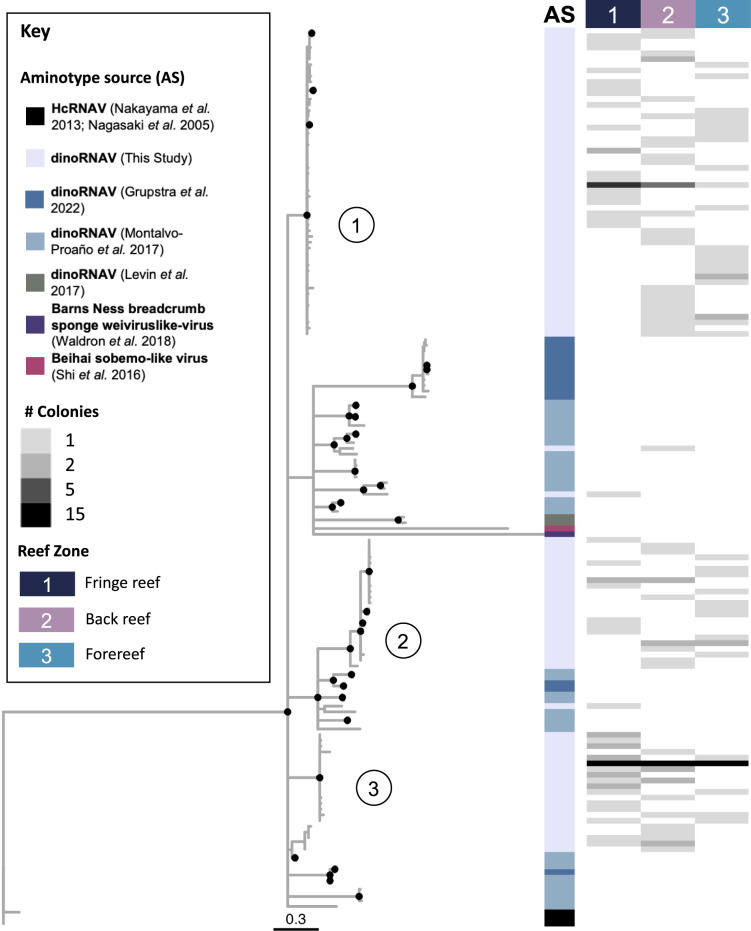
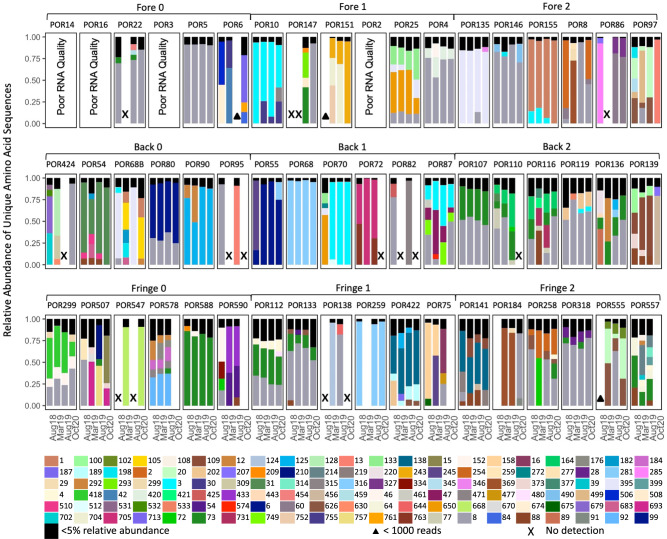
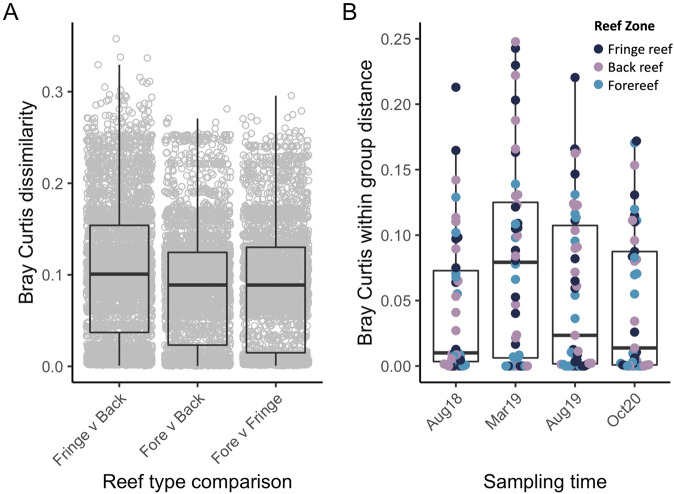
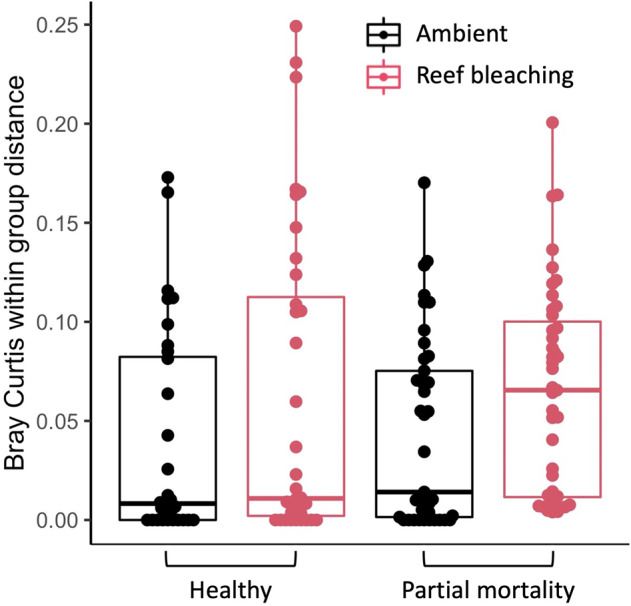
Viruses can affect coral health by infecting their symbiotic dinoflagellate partners (Symbiodiniaceae). Yet, viral dynamics in coral colonies exposed to environmental stress have not been studied at the reef scale, particularly within individual viral lineages. We sequenced the viral major capsid protein (mcp) gene of positive-sense single-stranded RNA viruses known to infect symbiotic dinoflagellates ('dinoRNAVs') to analyze their dynamics in the reef-building coral, Porites lobata. We repeatedly sampled 54 colonies harboring Cladocopium C15 dinoflagellates, across three environmentally distinct reef zones (fringing reef, back reef, and forereef) around the island of Moorea, French Polynesia over a 3-year period and spanning a reef-wide thermal stress event. By the end of the sampling period, 28% (5/18) of corals in the fringing reef experienced partial mortality versus 78% (14/18) of corals in the forereef. Over 90% (50/54) of colonies had detectable dinoRNAV infections. Reef zone influenced the composition and richness of viral mcp amino acid types ('aminotypes'), with the fringing reef containing the highest aminotype richness. The reef-wide thermal stress event significantly increased aminotype dispersion, and this pattern was strongest in the colonies that experienced partial mortality. These findings demonstrate that dinoRNAV infections respond to environmental fluctuations experienced in situ on reefs. Further, viral productivity will likely increase as ocean temperatures continue to rise, potentially impacting the foundational symbiosis underpinning coral reef ecosystems.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
