

Dynamics of inflammatory cytokine expression in bovine endometrial cells exposed to cow blood plasma small extracellular vesicles (sEV) may reflect high fertility
- PMID: 37012302
- PMCID: PMC10070242
- DOI: 10.1038/s41598-023-32045-1
Dynamics of inflammatory cytokine expression in bovine endometrial cells exposed to cow blood plasma small extracellular vesicles (sEV) may reflect high fertility
Abstract
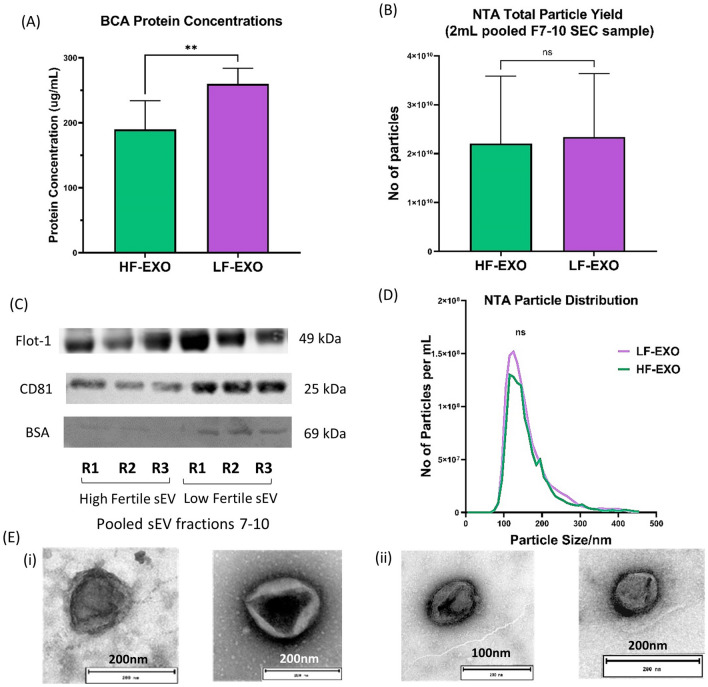
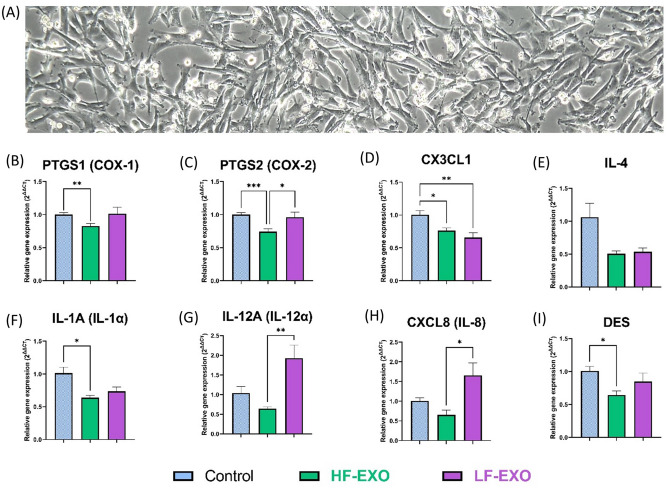
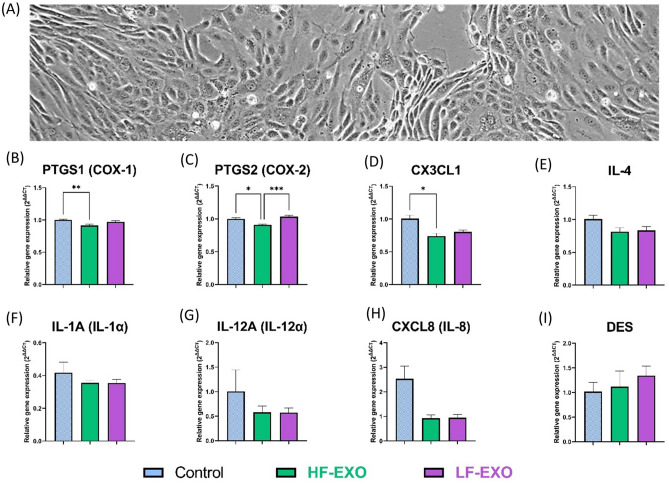
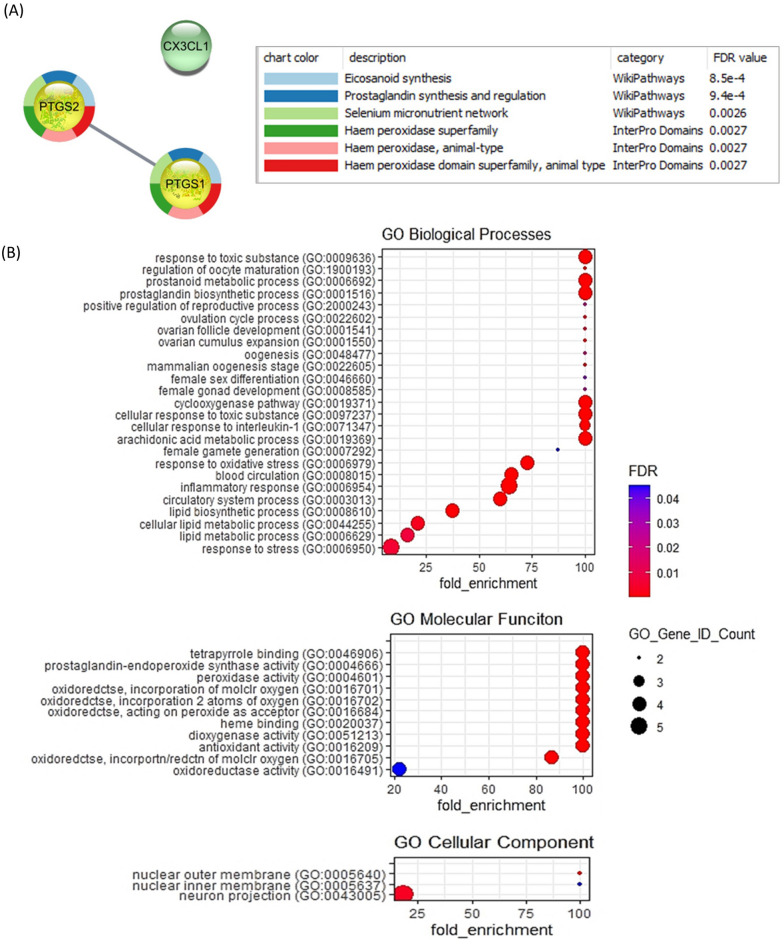
Aberrant inflammation in the endometrium impairs reproduction and leads to poor fertility. Small extracellular vesicles (sEV) are nanoparticles 30-200 nm in-size and contain transferable bioactive molecules that reflect the parent cell. Holstein-Friesian dairy cows with divergent genetic merit, high- (n = 10) and low-fertile (n = 10), were identified based on fertility breeding value (FBV), cow ovulation synchronization and postpartum anovulatory intervals (PPAI). In this study, we evaluated the effects of sEVs enriched from plasma of high-fertile (HF-EXO) and low-fertile (LF-EXO) dairy cows on inflammatory mediator expression by bovine endometrial epithelial (bEEL) and stromal (bCSC) cells. Exposure to HF-EXO in bCSC and bEEL cells yielded lower expression of PTGS1 and PTGS2 compared to the control. In bCSC cells exposed to HF-EXO, pro-inflammatory cytokine IL1-α was downregulated compared to the untreated control, IL-12α and IL-8 were downregulated compared to the LF-EXO treatment. Our findings demonstrate that sEVs interact with both endometrial epithelial and stromal cells to initiate differential gene expression, specifically genes relate to inflammation. Therefore, even subtle changes on the inflammatory gene cascade in the endometrium via sEV may affect reproductive performance and/or outcomes. Further, sEV from high-fertile animals acts in a unique direction to deactivate prostaglandin synthases in both bCSC and bEEL cells and deactivate pro-inflammatory cytokines in the endometrial stroma. The results suggest that circulating sEV may serve as a potential biomarker of fertility.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
