

Neural dynamics underlying associative learning in the dorsal and ventral hippocampus
- PMID: 37012382
- PMCID: PMC10448873
- DOI: 10.1038/s41593-023-01296-6
Neural dynamics underlying associative learning in the dorsal and ventral hippocampus
Abstract
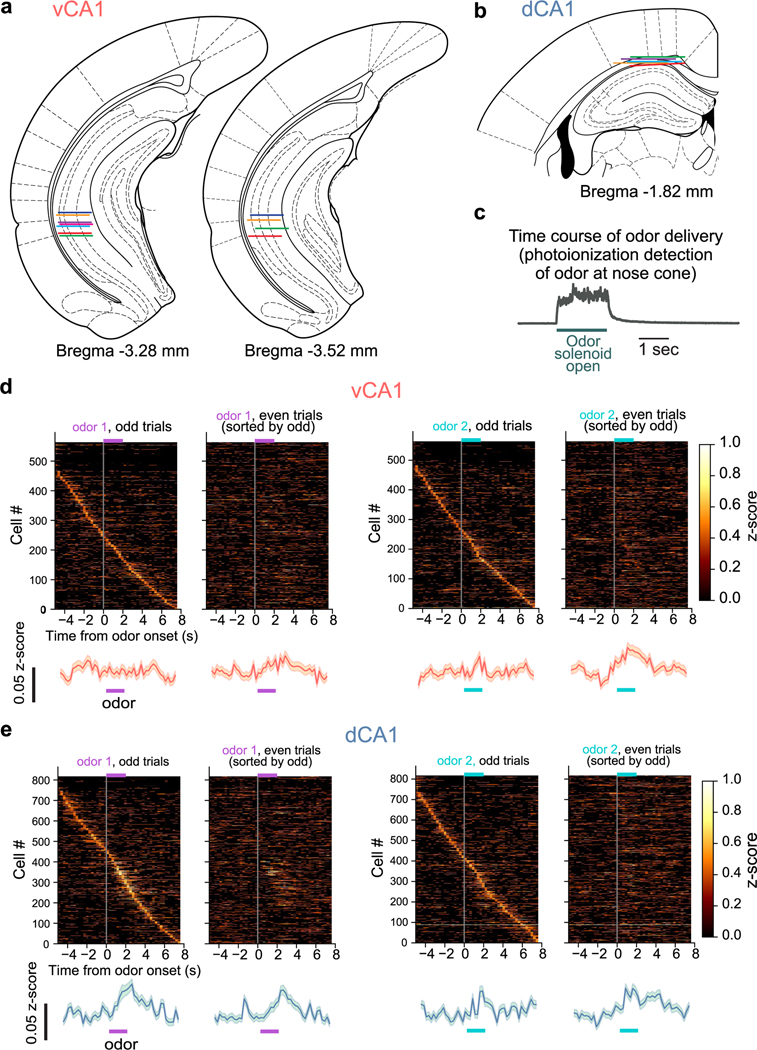
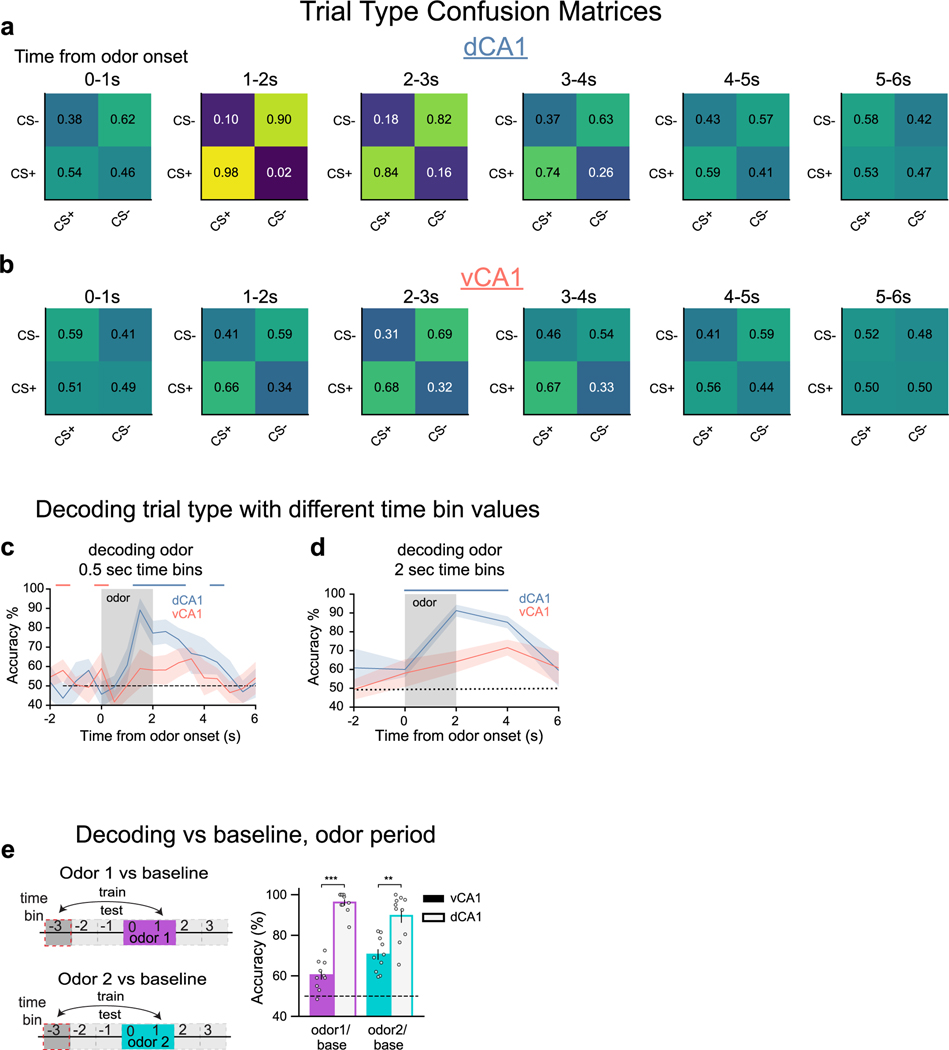
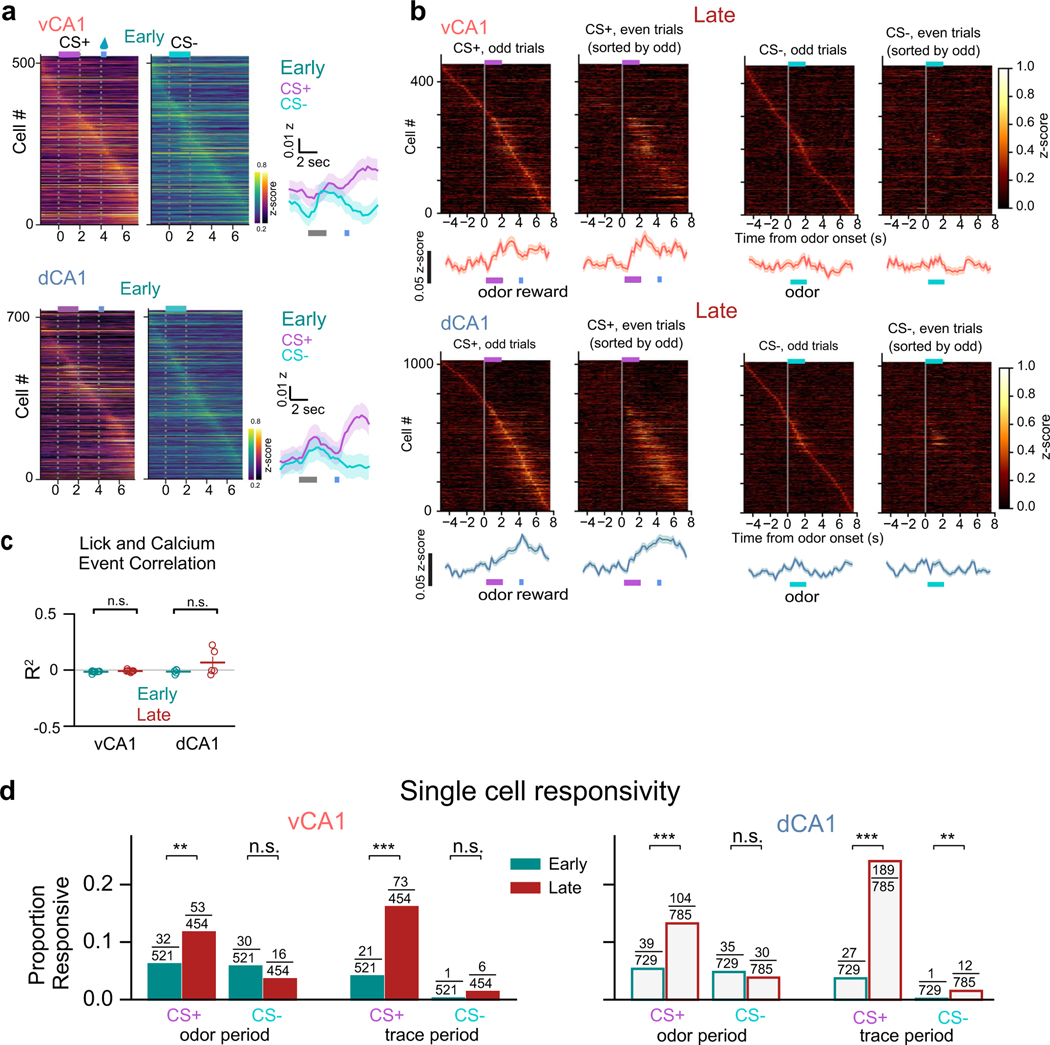
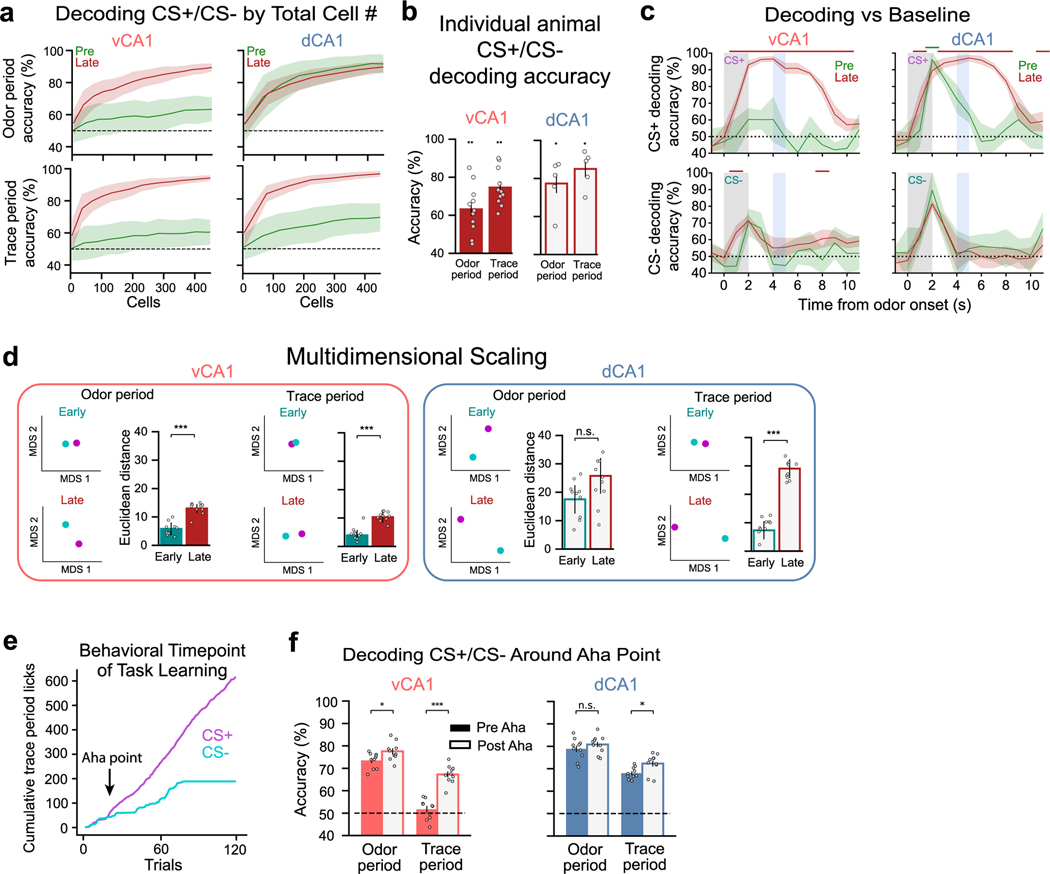
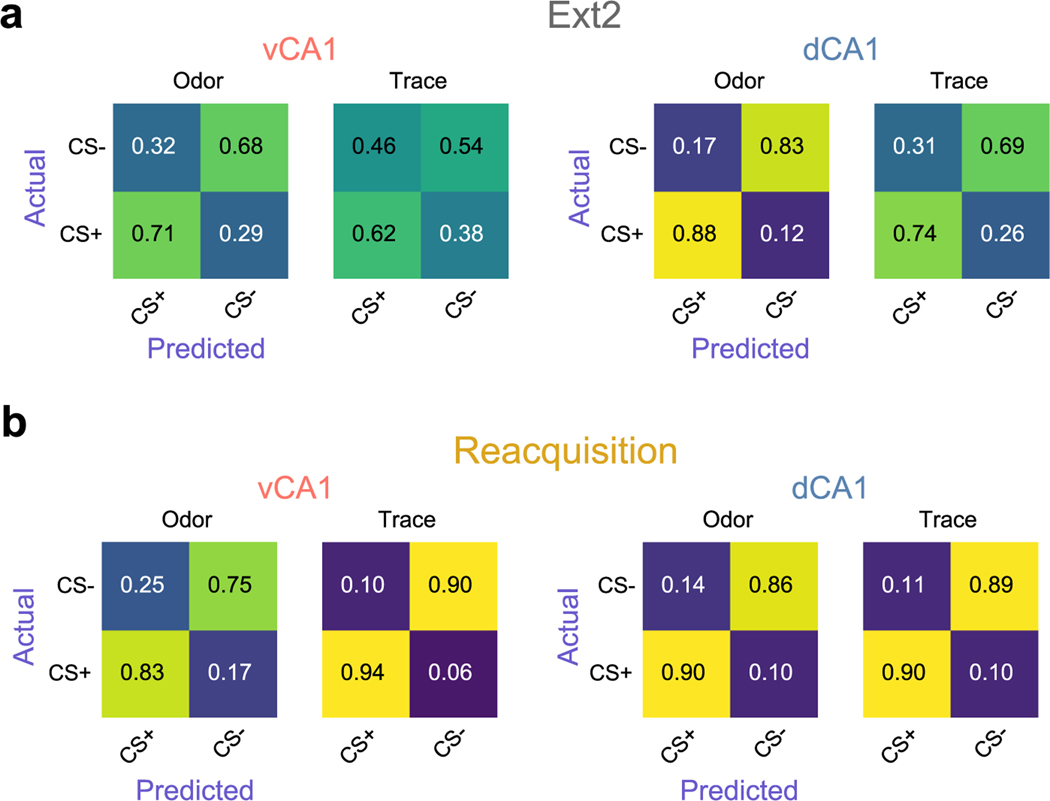
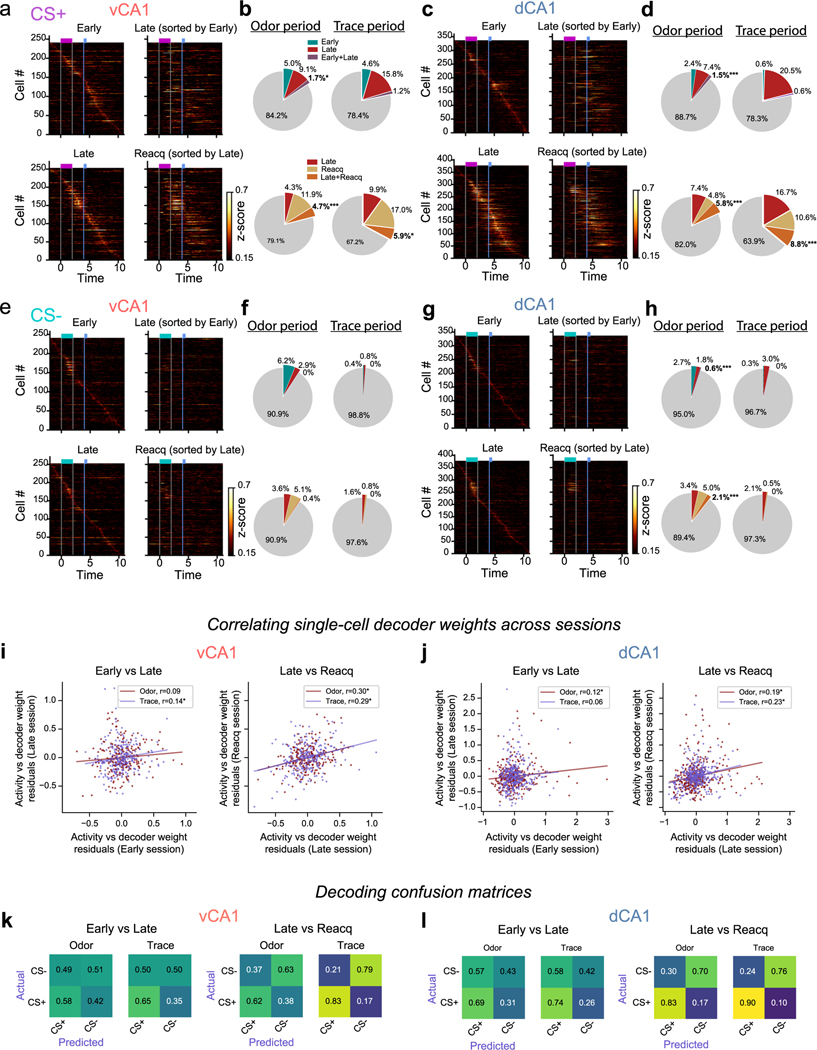
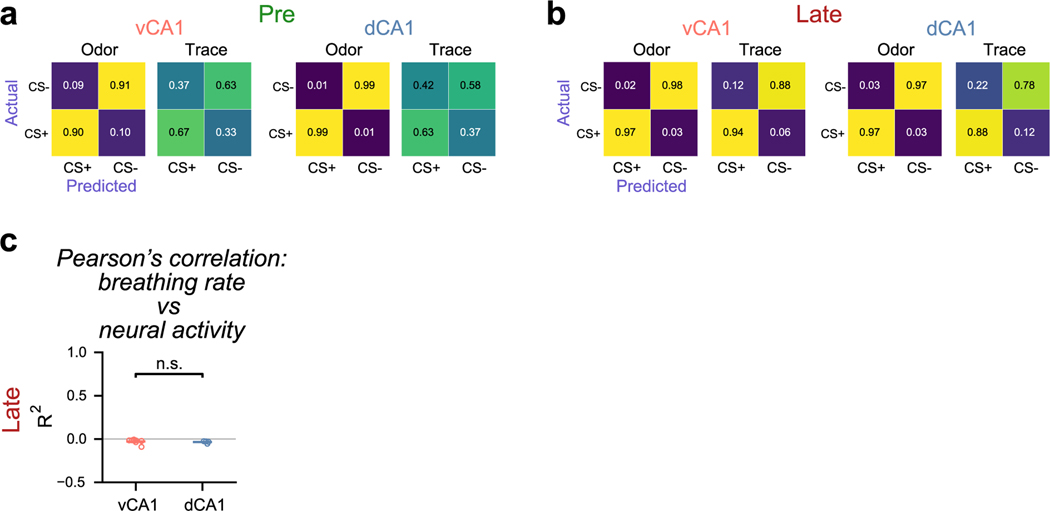
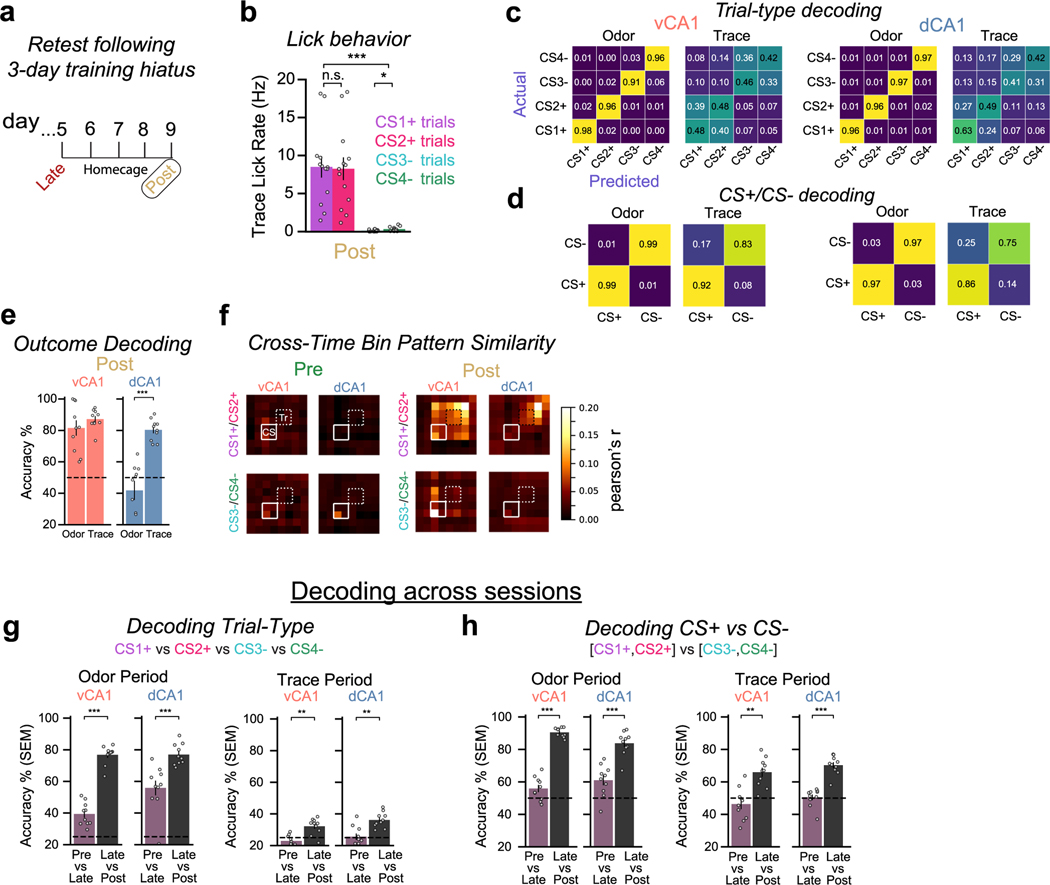
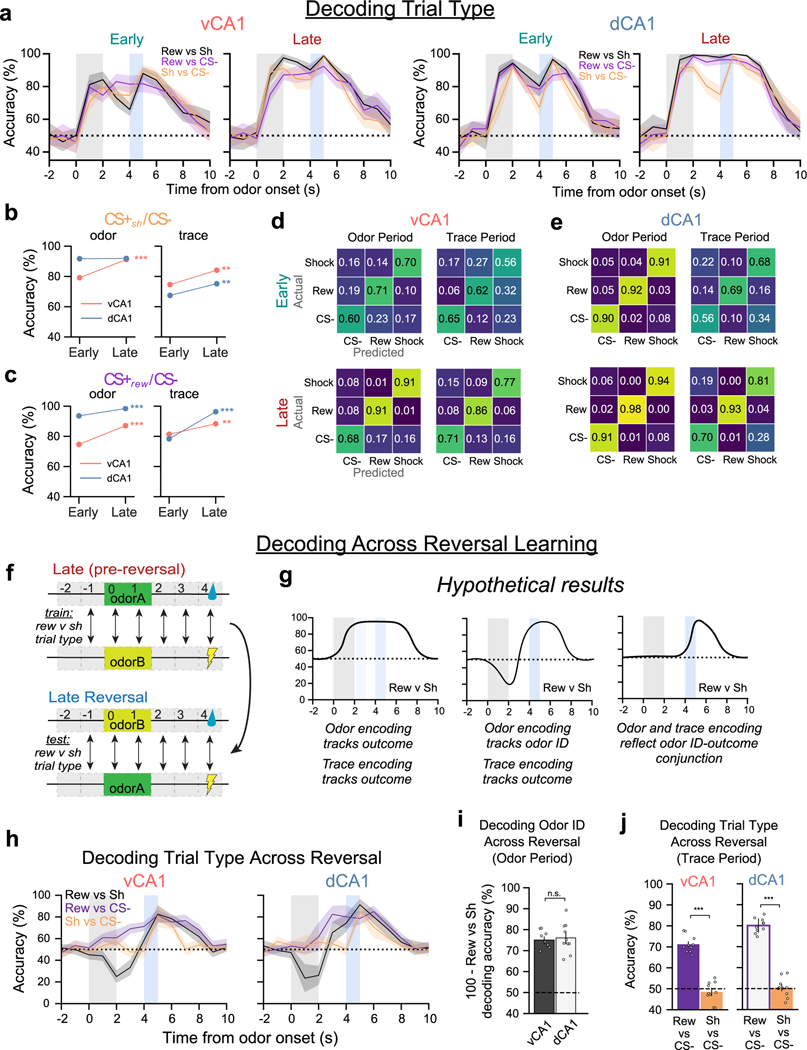
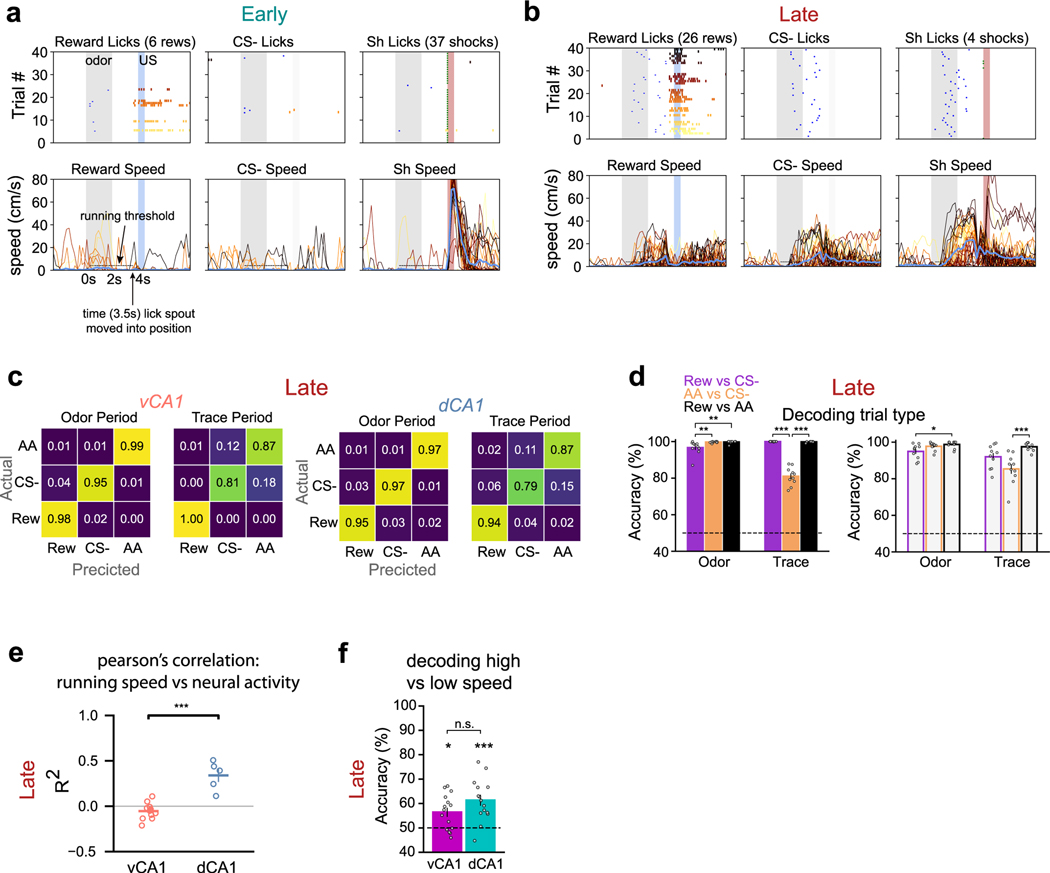
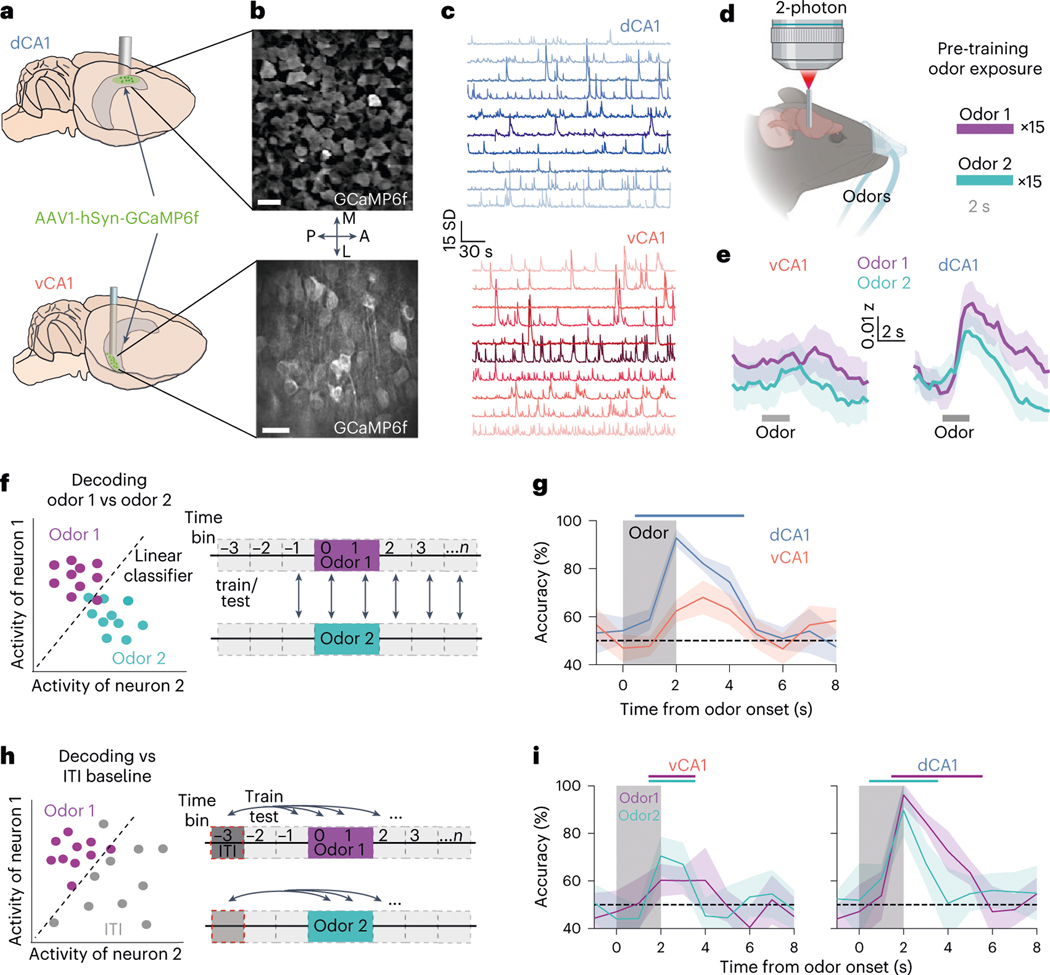
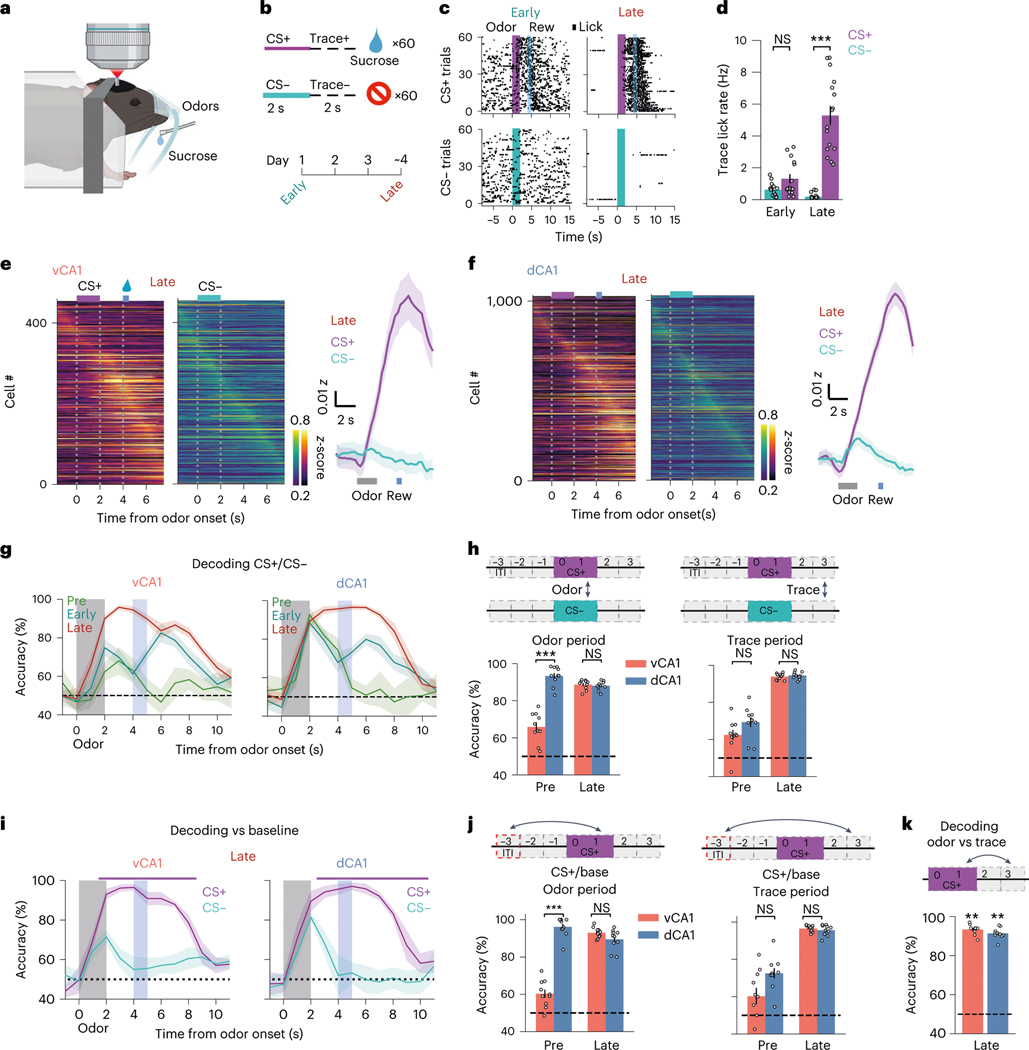
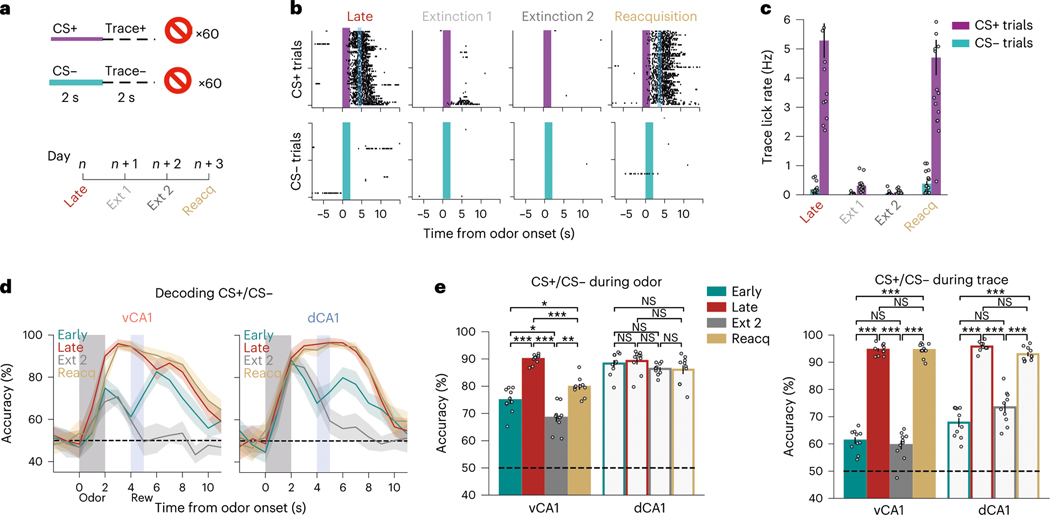
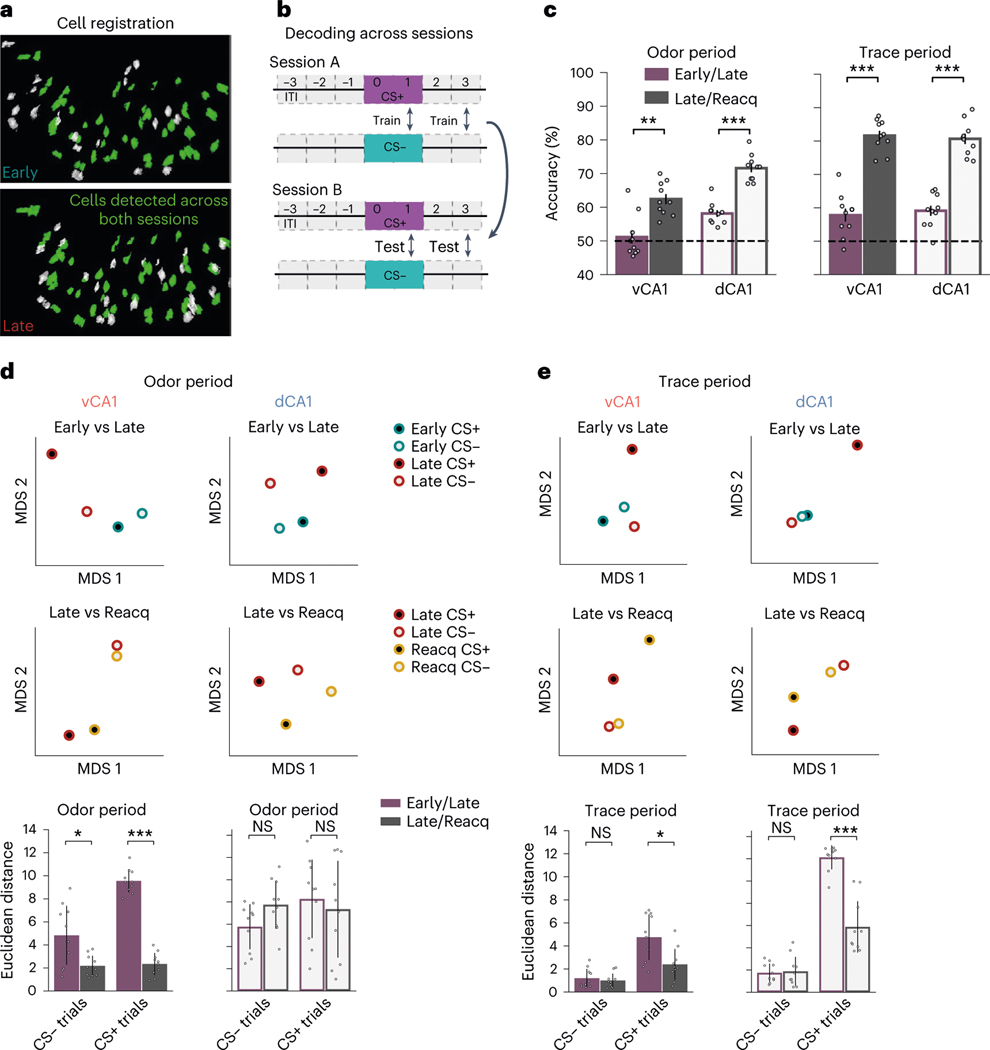
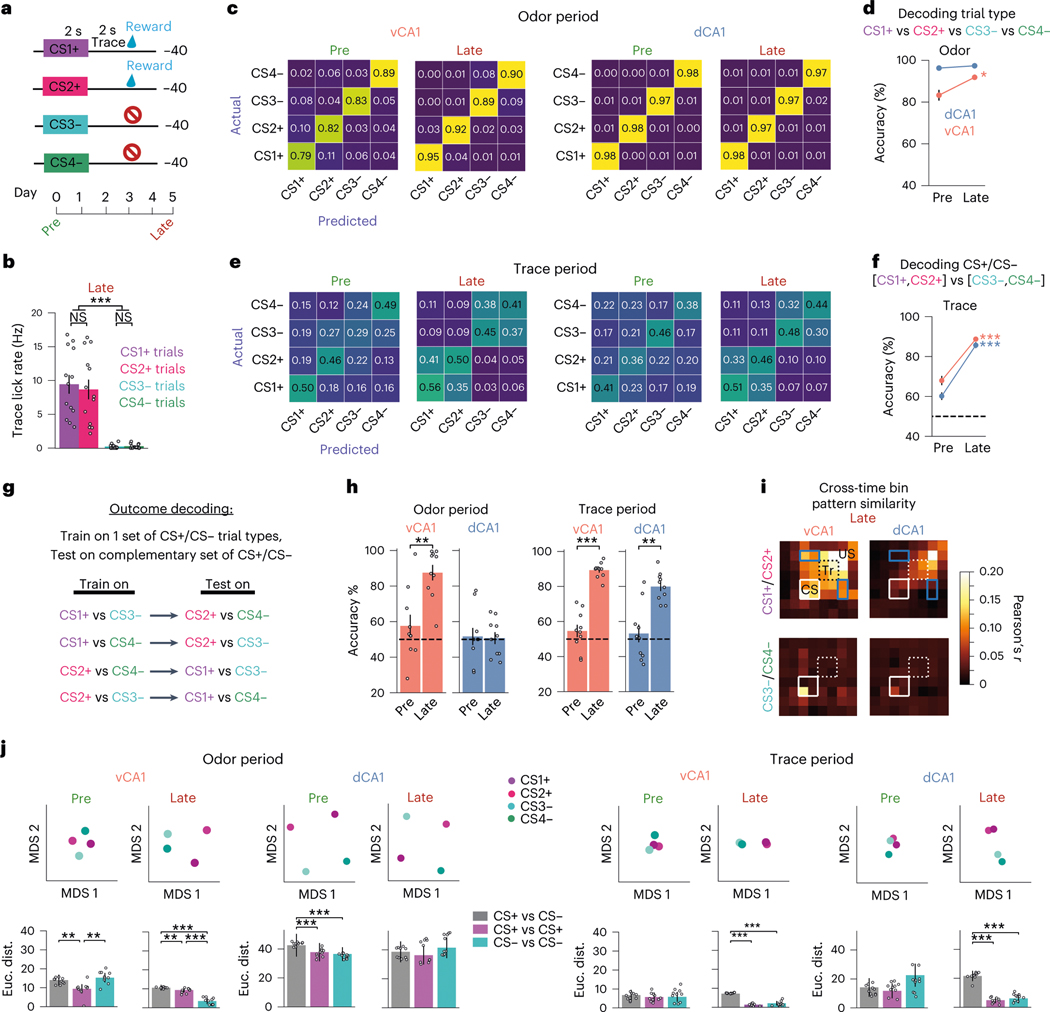
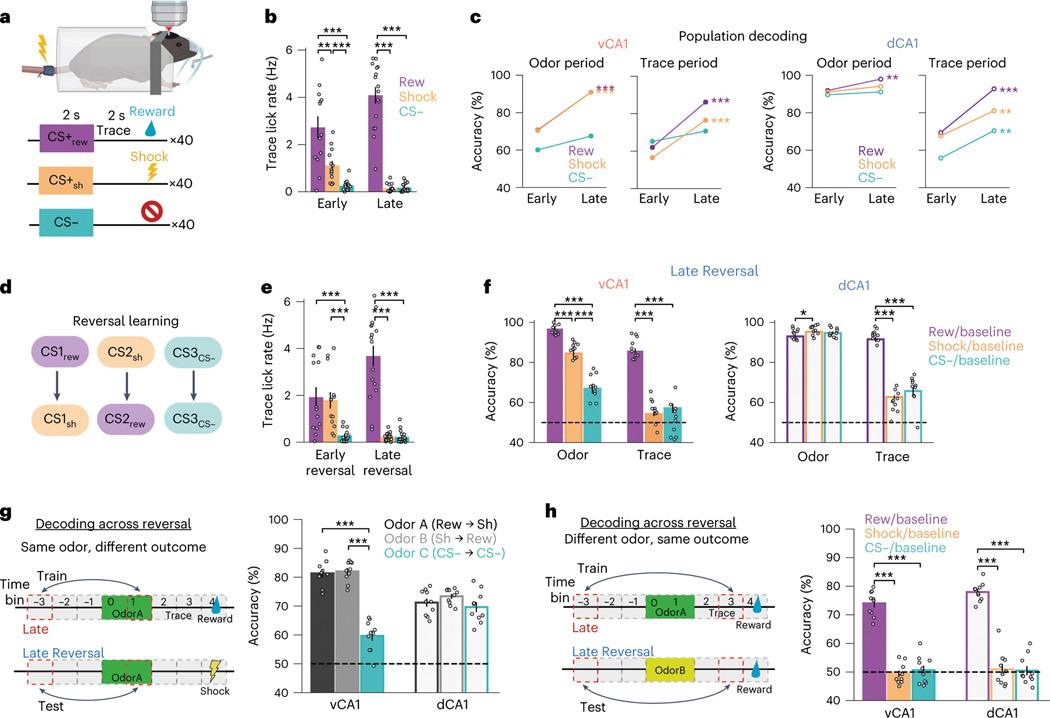
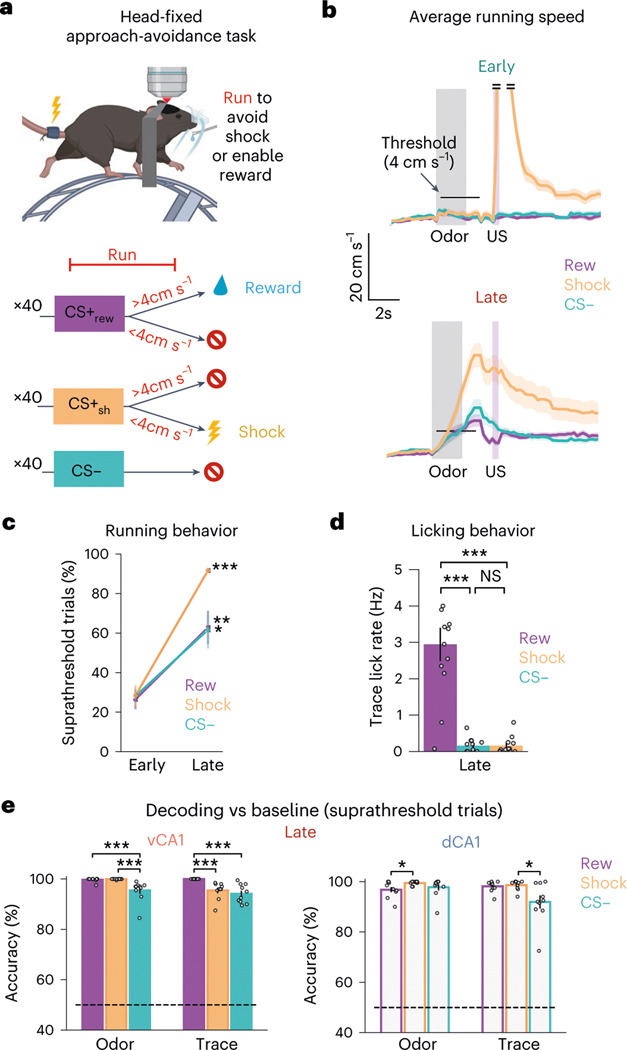
Animals associate cues with outcomes and update these associations as new information is presented. This requires the hippocampus, yet how hippocampal neurons track changes in cue-outcome associations remains unclear. Using two-photon calcium imaging, we tracked the same dCA1 and vCA1 neurons across days to determine how responses evolve across phases of odor-outcome learning. Initially, odors elicited robust responses in dCA1, whereas, in vCA1, odor responses primarily emerged after learning and embedded information about the paired outcome. Population activity in both regions rapidly reorganized with learning and then stabilized, storing learned odor representations for days, even after extinction or pairing with a different outcome. Additionally, we found stable, robust signals across CA1 when mice anticipated outcomes under behavioral control but not when mice anticipated an inescapable aversive outcome. These results show how the hippocampus encodes, stores and updates learned associations and illuminates the unique contributions of dorsal and ventral hippocampus.
© 2023. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Strange BA, Witter MP, Lein ES & Moser EI Functional organization of the hippocampal longitudinal axis. Nat. Rev. Neurosci 15, 655–669 (2014). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
