

Rewiring of the 3D genome during acquisition of carboplatin resistance in a triple-negative breast cancer patient-derived xenograft
- PMID: 37012431
- PMCID: PMC10070455
- DOI: 10.1038/s41598-023-32568-7
Rewiring of the 3D genome during acquisition of carboplatin resistance in a triple-negative breast cancer patient-derived xenograft
Erratum in
-
Publisher Correction: Rewiring of the 3D genome during acquisition of carboplatin resistance in a triple-negative breast cancer patient-derived xenograft.Sci Rep. 2023 May 9;13(1):7522. doi: 10.1038/s41598-023-33284-y. Sci Rep. 2023. PMID: 37160896 Free PMC article. No abstract available.
-
Author Correction: Rewiring of the 3D genome during acquisition of carboplatin resistance in a triple-negative breast cancer patient-derived xenograft.Sci Rep. 2024 Dec 17;14(1):30519. doi: 10.1038/s41598-024-82374-y. Sci Rep. 2024. PMID: 39690151 Free PMC article. No abstract available.
Abstract
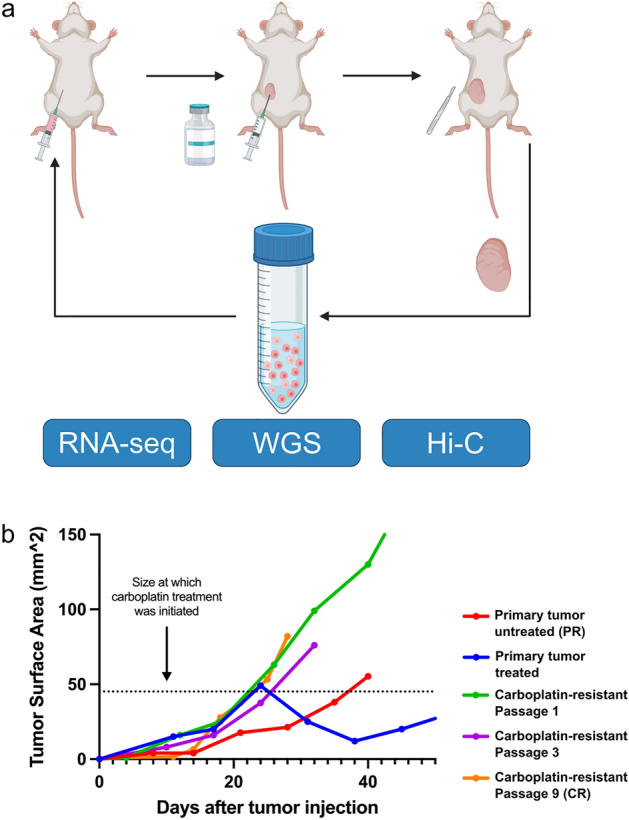
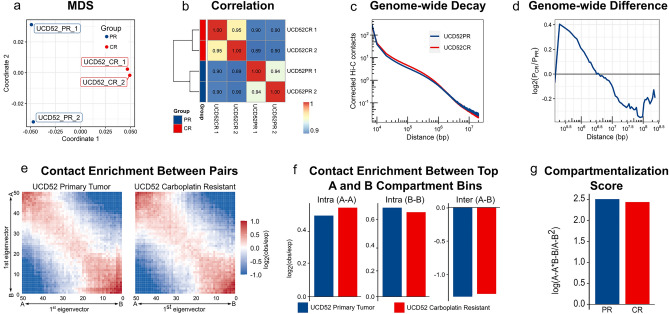
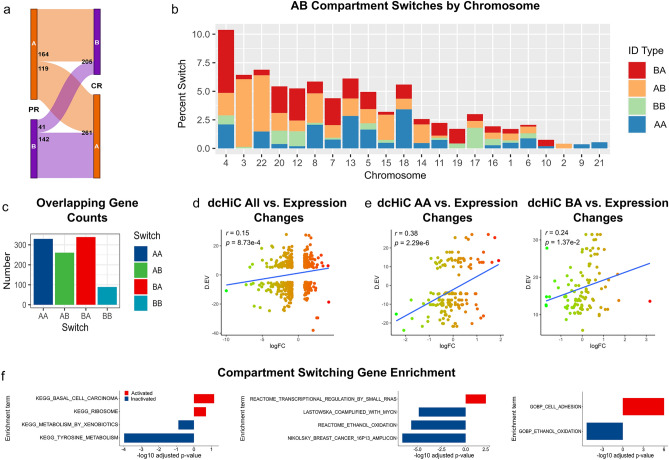
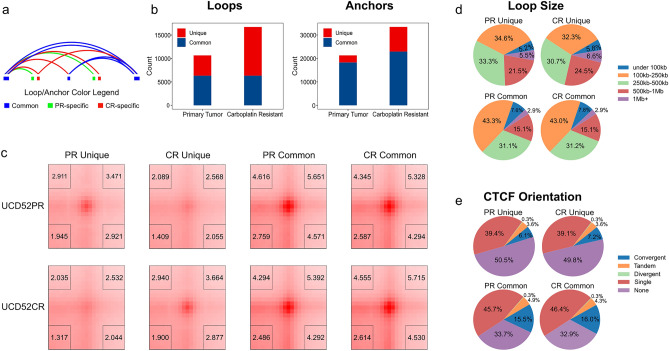
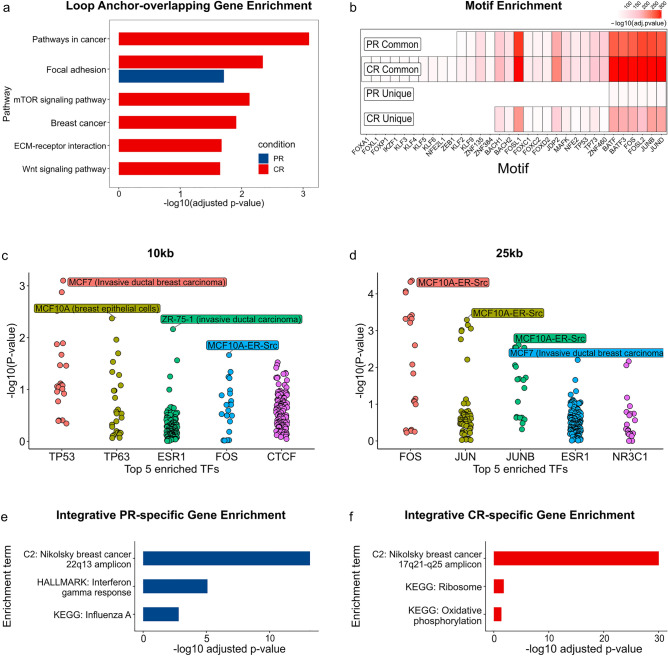
Changes in the three-dimensional (3D) structure of the genome are an emerging hallmark of cancer. Cancer-associated copy number variants and single nucleotide polymorphisms promote rewiring of chromatin loops, disruption of topologically associating domains (TADs), active/inactive chromatin state switching, leading to oncogene expression and silencing of tumor suppressors. However, little is known about 3D changes during cancer progression to a chemotherapy-resistant state. We integrated chromatin conformation capture (Hi-C), RNA-seq, and whole-genome sequencing obtained from triple-negative breast cancer patient-derived xenograft primary tumors (UCD52) and carboplatin-resistant samples and found increased short-range (< 2 Mb) interactions, chromatin looping, formation of TAD, chromatin state switching into a more active state, and amplification of ATP-binding cassette transporters. Transcriptome changes suggested the role of long-noncoding RNAs in carboplatin resistance. Rewiring of the 3D genome was associated with TP53, TP63, BATF, FOS-JUN family of transcription factors and led to activation of aggressiveness-, metastasis- and other cancer-related pathways. Integrative analysis highlighted increased ribosome biogenesis and oxidative phosphorylation, suggesting the role of mitochondrial energy metabolism. Our results suggest that 3D genome remodeling may be a key mechanism underlying carboplatin resistance.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
