

Toll-Like Receptor 3 Mediates Aortic Stenosis Through a Conserved Mechanism of Calcification
- PMID: 37013819
- PMCID: PMC10192061
- DOI: 10.1161/CIRCULATIONAHA.122.063481
Toll-Like Receptor 3 Mediates Aortic Stenosis Through a Conserved Mechanism of Calcification
Abstract
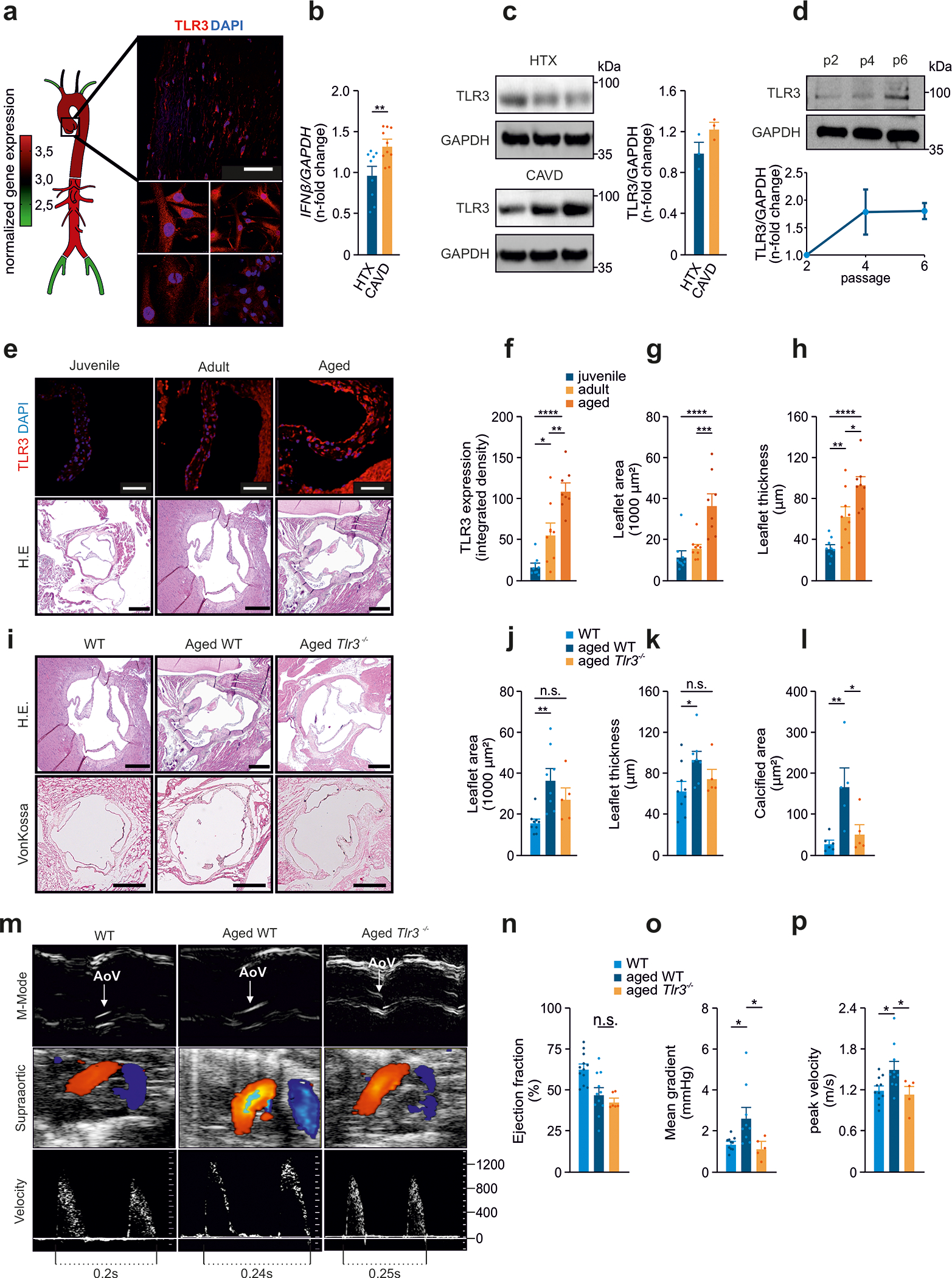
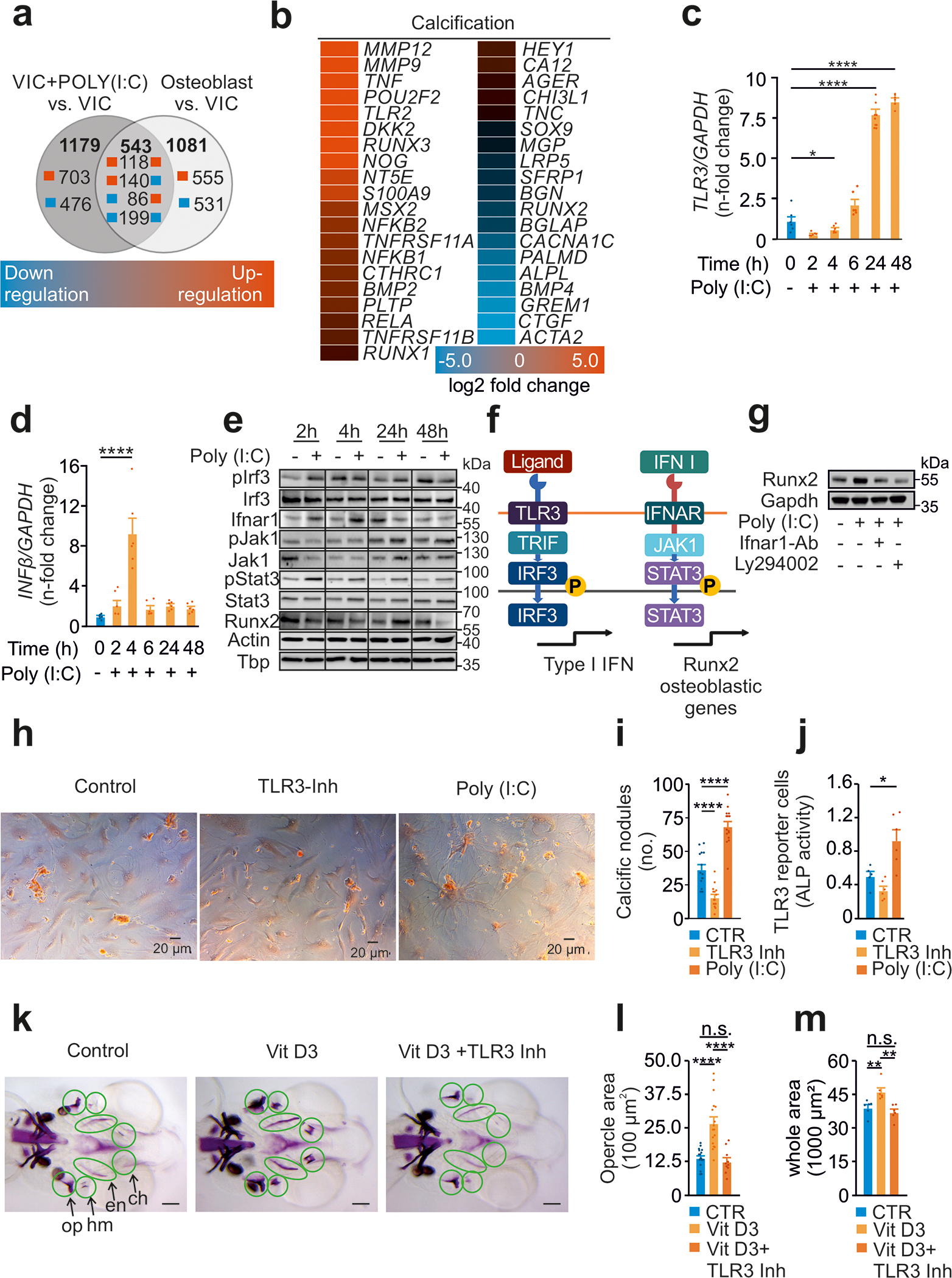
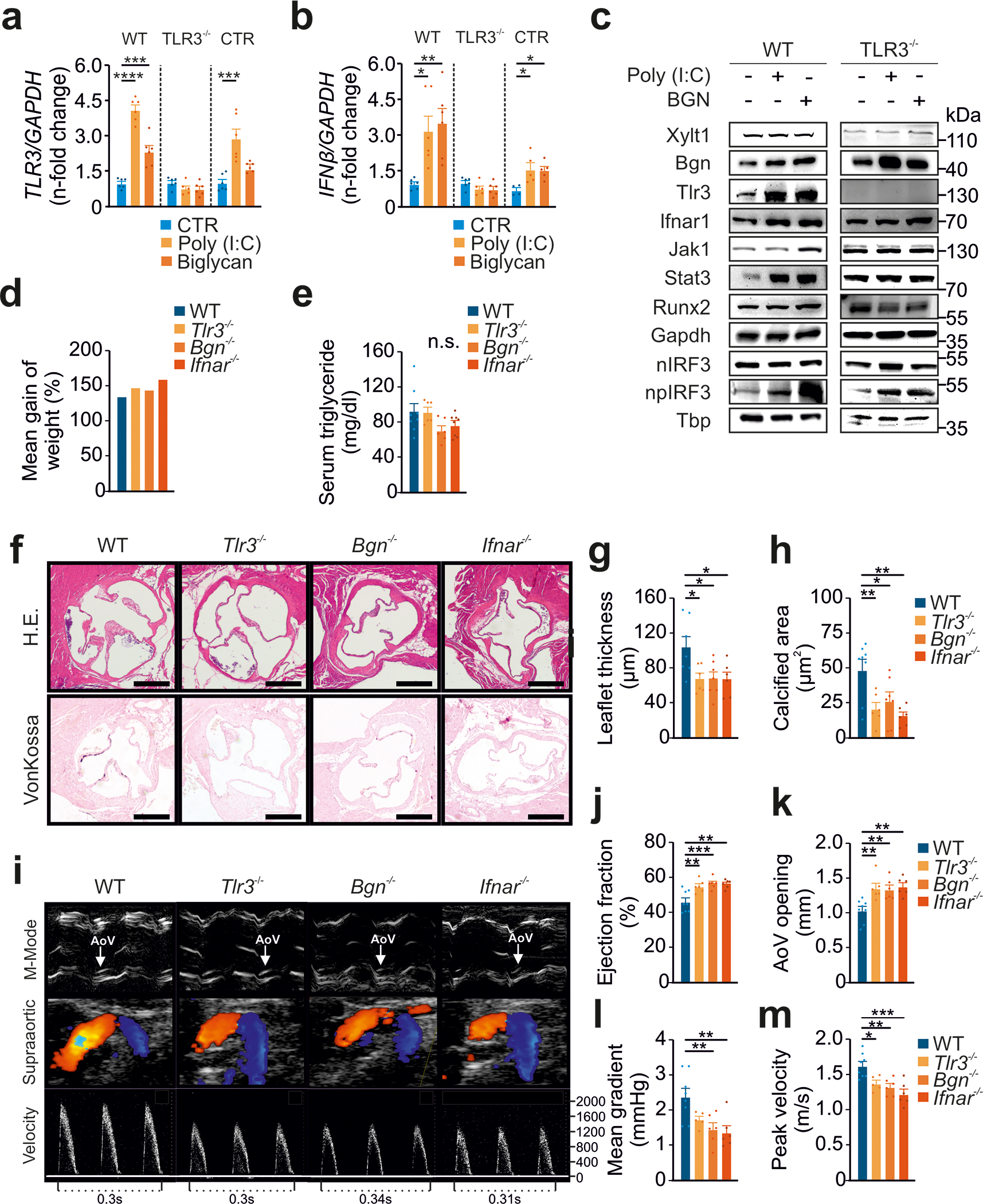
Background: Calcific aortic valve disease (CAVD) is characterized by a phenotypic switch of valvular interstitial cells to bone-forming cells. Toll-like receptors (TLRs) are evolutionarily conserved pattern recognition receptors at the interface between innate immunity and tissue repair. Type I interferons (IFNs) are not only crucial for an adequate antiviral response but also implicated in bone formation. We hypothesized that the accumulation of endogenous TLR3 ligands in the valvular leaflets may promote the generation of osteoblast-like cells through enhanced type I IFN signaling.
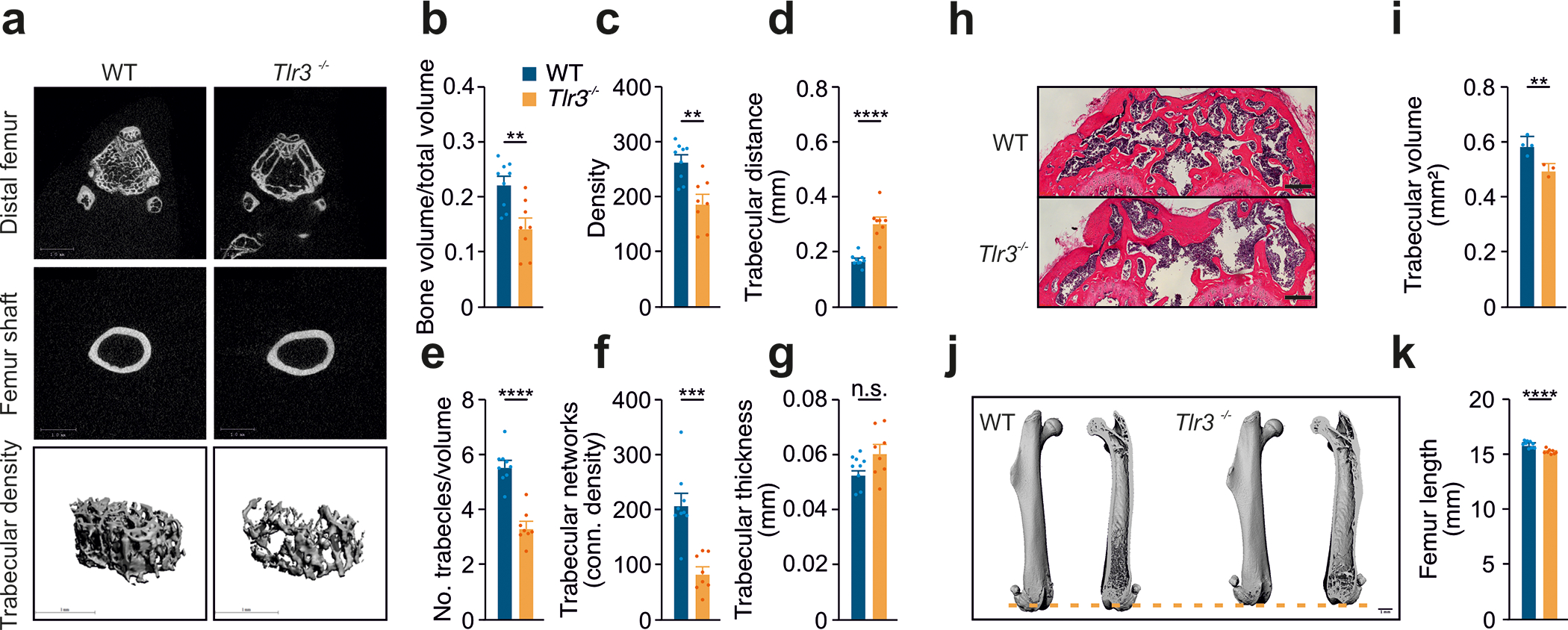
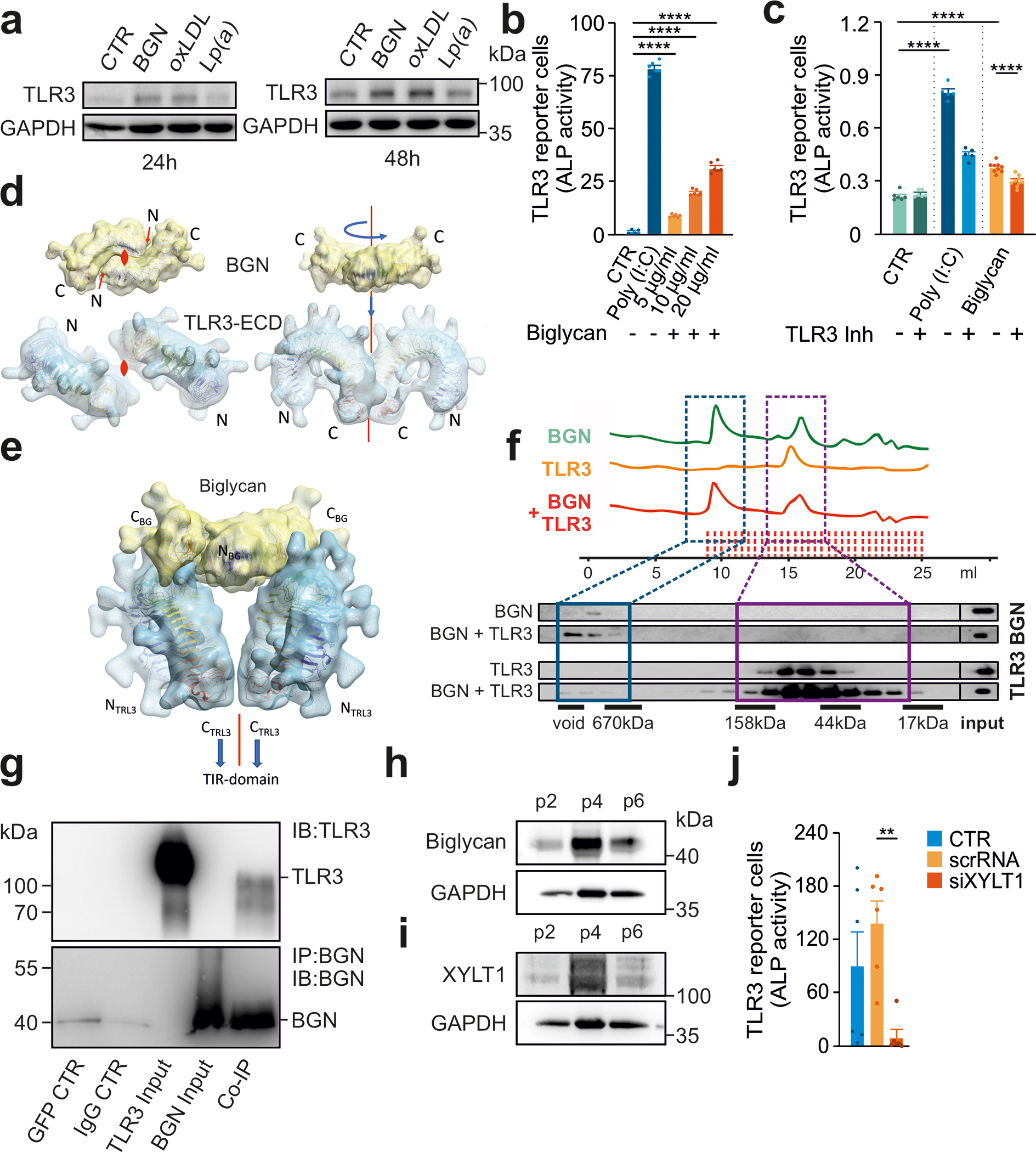
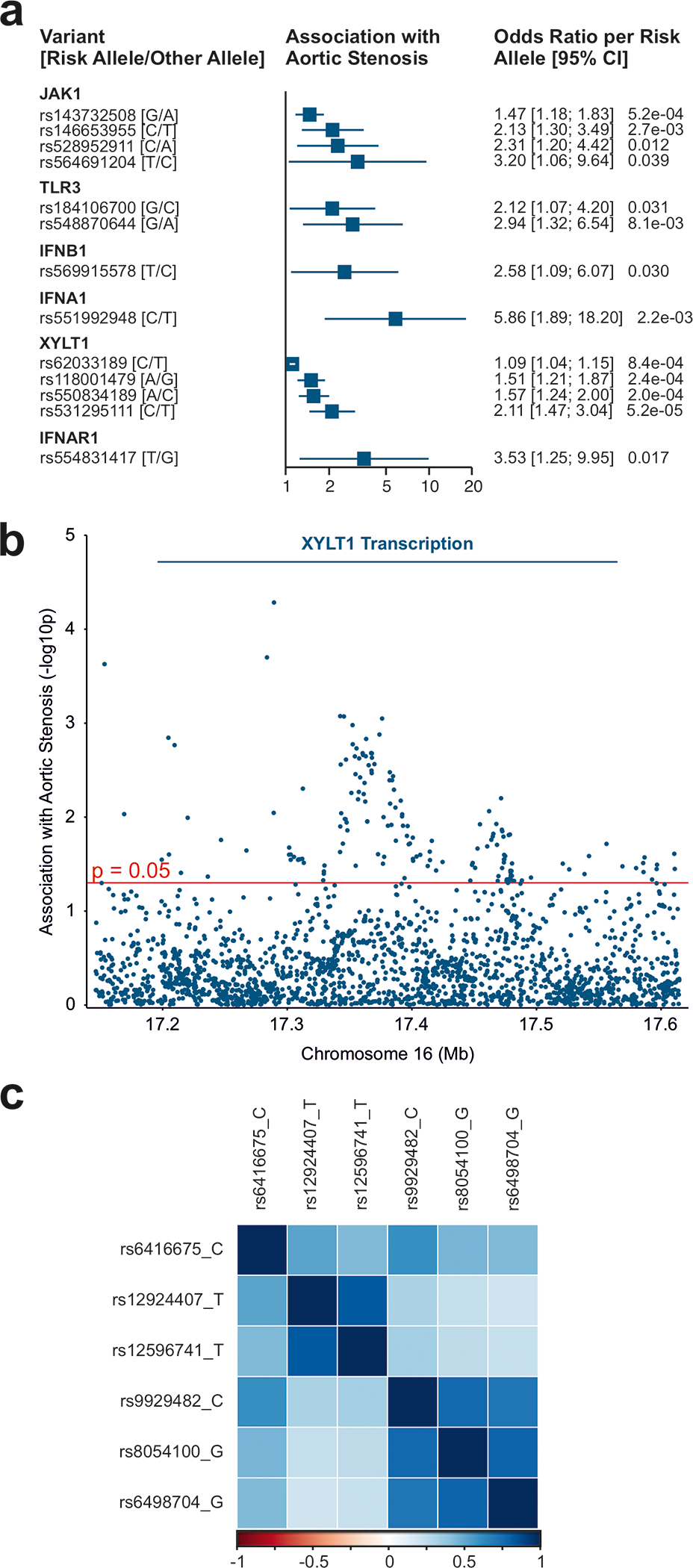
Methods: Human valvular interstitial cells isolated from aortic valves were challenged with mechanical strain or synthetic TLR3 agonists and analyzed for bone formation, gene expression profiles, and IFN signaling pathways. Different inhibitors were used to delineate the engaged signaling pathways. Moreover, we screened a variety of potential lipids and proteoglycans known to accumulate in CAVD lesions as potential TLR3 ligands. Ligand-receptor interactions were characterized by in silico modeling and verified through immunoprecipitation experiments. Biglycan (Bgn), Tlr3, and IFN-α/β receptor alpha chain (Ifnar1)-deficient mice and a specific zebrafish model were used to study the implication of the biglycan (BGN)-TLR3-IFN axis in both CAVD and bone formation in vivo. Two large-scale cohorts (GERA [Genetic Epidemiology Research on Adult Health and Aging], n=55 192 with 3469 aortic stenosis cases; UK Biobank, n=257 231 with 2213 aortic stenosis cases) were examined for genetic variation at genes implicated in BGN-TLR3-IFN signaling associating with CAVD in humans.
Results: Here, we identify TLR3 as a central molecular regulator of calcification in valvular interstitial cells and unravel BGN as a new endogenous agonist of TLR3. Posttranslational BGN maturation by xylosyltransferase 1 (XYLT1) is required for TLR3 activation. Moreover, BGN induces the transdifferentiation of valvular interstitial cells into bone-forming osteoblasts through the TLR3-dependent induction of type I IFNs. It is intriguing that Bgn-/-, Tlr3-/-, and Ifnar1-/- mice are protected against CAVD and display impaired bone formation. Meta-analysis of 2 large-scale cohorts with >300 000 individuals reveals that genetic variation at loci relevant to the XYLT1-BGN-TLR3-interferon-α/β receptor alpha chain (IFNAR) 1 pathway is associated with CAVD in humans.
Conclusions: This study identifies the BGN-TLR3-IFNAR1 axis as an evolutionarily conserved pathway governing calcification of the aortic valve and reveals a potential therapeutic target to prevent CAVD.
Keywords: Toll-like receptor 3; aortic valve disease; biglycan; extracellular matrix; osteogenesis; proteins.
Conflict of interest statement
Figures
References
-
- Rajamannan NM, Evans FJ, Aikawa E, Grande-Allen KJ, Demer LL, Heistad DD, Simmons CA, Masters KS, Mathieu P, O’Brien KD, et al. Calcific aortic valve disease: not simply a degenerative process: A review and agenda for research from the National Heart and Lung and Blood Institute Aortic Stenosis Working Group. Executive summary: Calcific aortic valve disease-2011 update. Circulation. 2011;124:1783–1791. doi: 10.1161/CIRCULATIONAHA.110.006767 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
