

Accumulation of α-synuclein mediates podocyte injury in Fabry nephropathy
- PMID: 37014703
- PMCID: PMC10232004
- DOI: 10.1172/JCI157782
Accumulation of α-synuclein mediates podocyte injury in Fabry nephropathy
Abstract
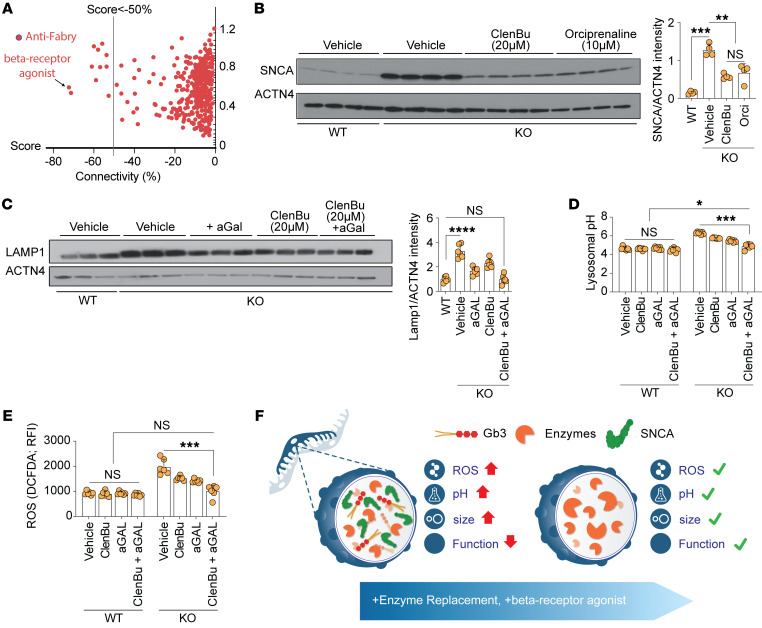
Current therapies for Fabry disease are based on reversing intracellular accumulation of globotriaosylceramide (Gb3) by enzyme replacement therapy (ERT) or chaperone-mediated stabilization of the defective enzyme, thereby alleviating lysosomal dysfunction. However, their effect in the reversal of end-organ damage, like kidney injury and chronic kidney disease, remains unclear. In this study, ultrastructural analysis of serial human kidney biopsies showed that long-term use of ERT reduced Gb3 accumulation in podocytes but did not reverse podocyte injury. Then, a CRISPR/Cas9-mediated α-galactosidase knockout podocyte cell line confirmed ERT-mediated reversal of Gb3 accumulation without resolution of lysosomal dysfunction. Transcriptome-based connectivity mapping and SILAC-based quantitative proteomics identified α-synuclein (SNCA) accumulation as a key event mediating podocyte injury. Genetic and pharmacological inhibition of SNCA improved lysosomal structure and function in Fabry podocytes, exceeding the benefits of ERT. Together, this work reconceptualizes Fabry-associated cell injury beyond Gb3 accumulation, and introduces SNCA modulation as a potential intervention, especially for patients with Fabry nephropathy.
Keywords: Chronic kidney disease; Drug therapy; Genetic diseases; Genetics; Nephrology.
Figures
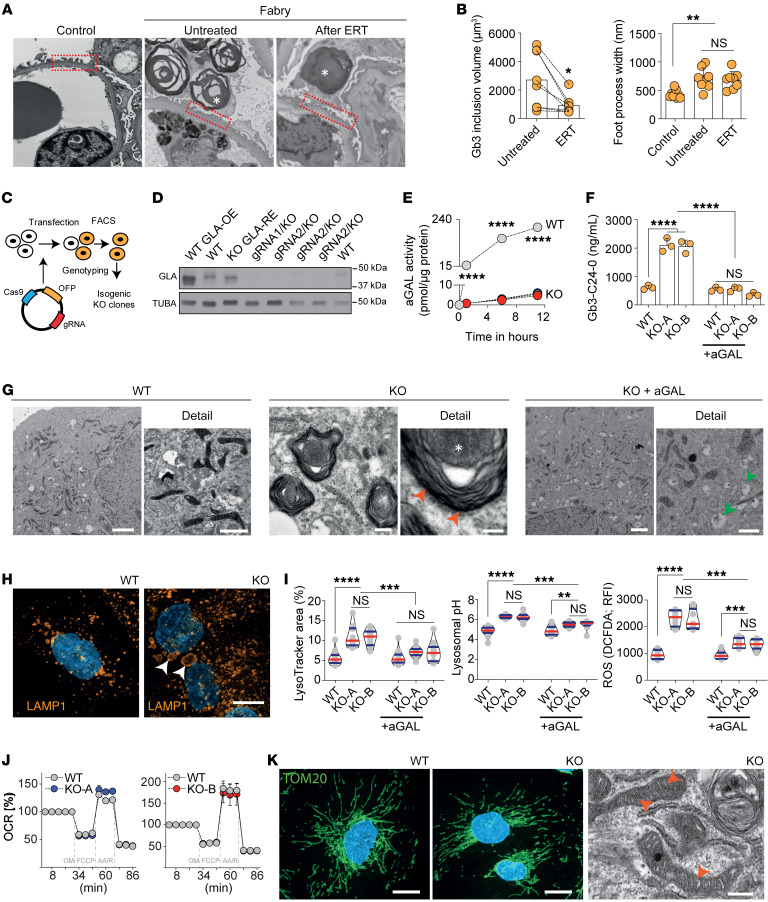
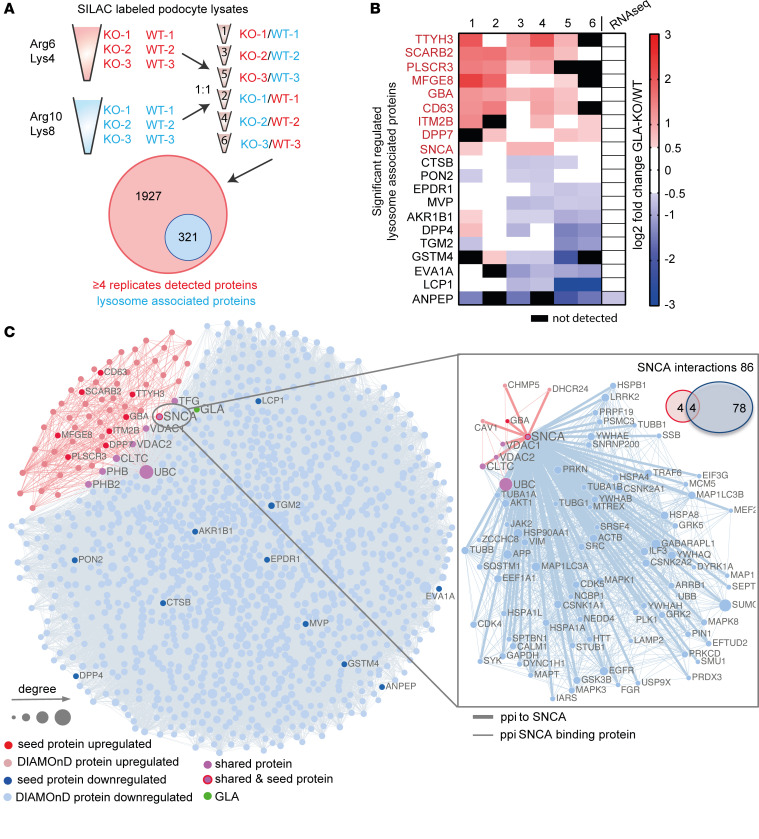
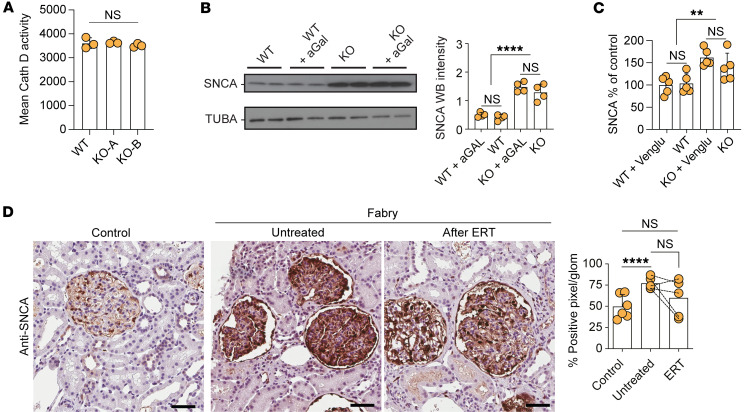
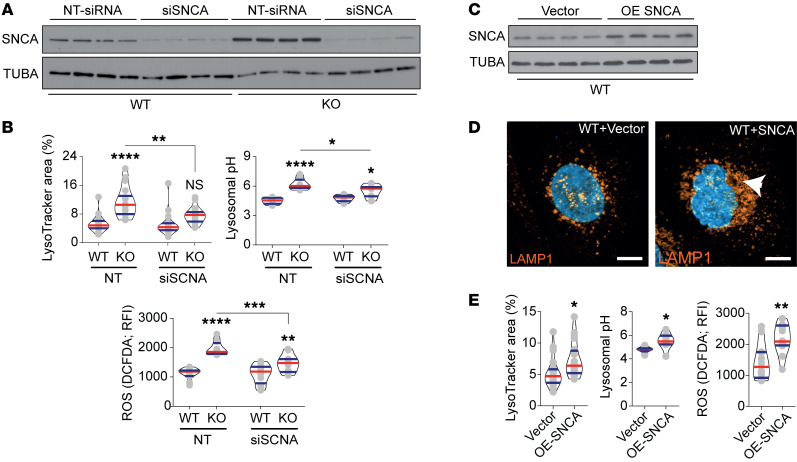
Comment in
-
Key role of α-synuclein in Fabry nephropathy.Nat Rev Nephrol. 2023 Jun;19(6):359. doi: 10.1038/s41581-023-00723-y. Nat Rev Nephrol. 2023. PMID: 37130928 No abstract available.
References
-
- Anderson W. A case of “angeio-keratoma. ” Br J Dermatol. 1898;10(4):113–117. doi: 10.1111/j.1365-2133.1898.tb16317.x. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
