

Conditional permeabilization of the P. falciparum plasma membrane in infected cells links cation influx to reduced membrane integrity
- PMID: 37014920
- PMCID: PMC10072447
- DOI: 10.1371/journal.pone.0283776
Conditional permeabilization of the P. falciparum plasma membrane in infected cells links cation influx to reduced membrane integrity
Abstract
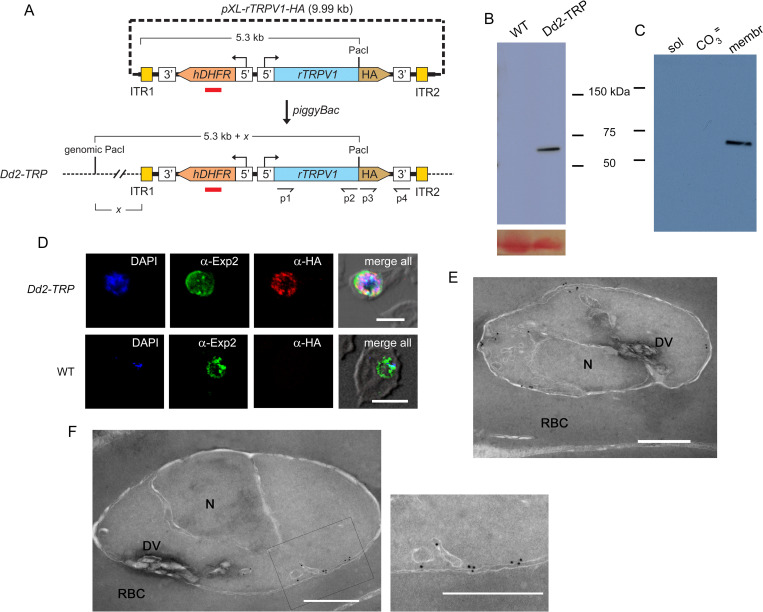
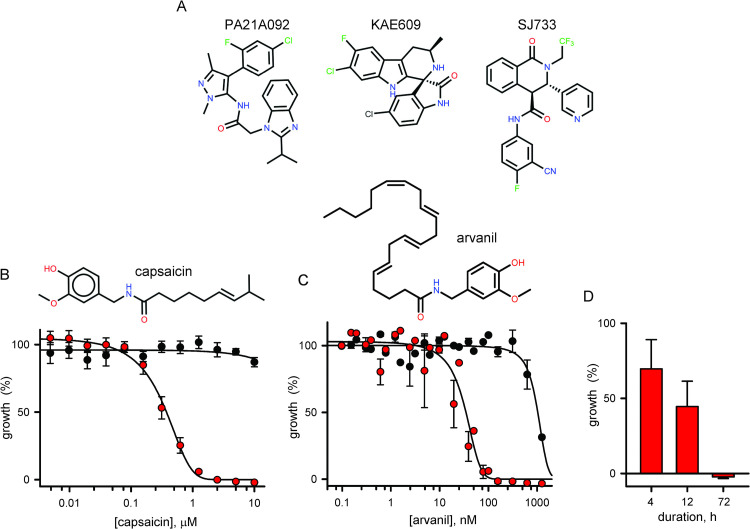
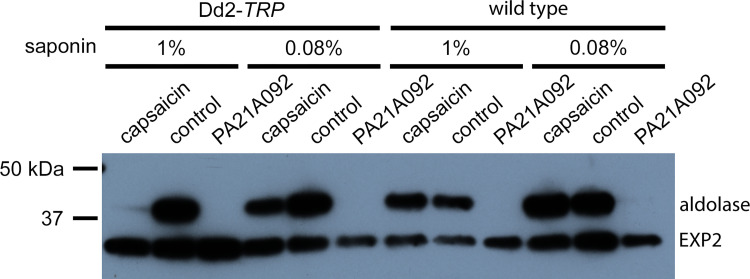
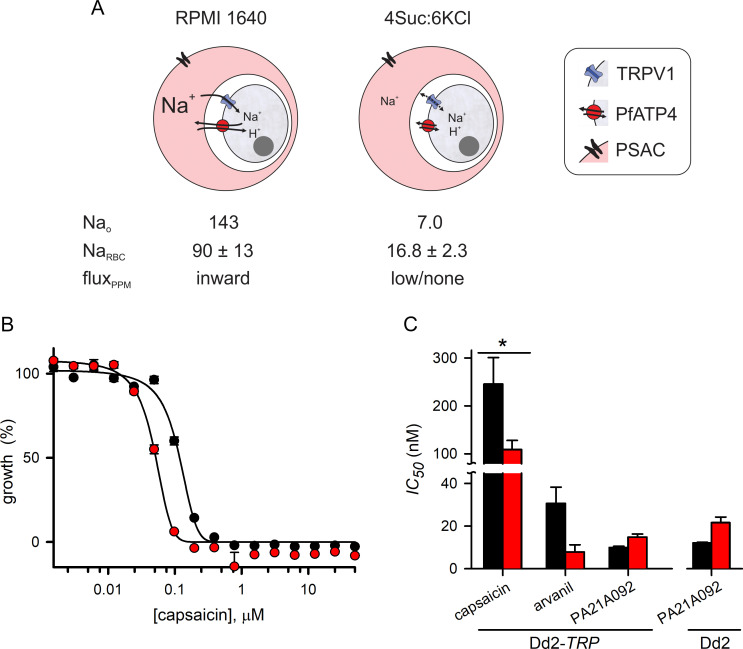
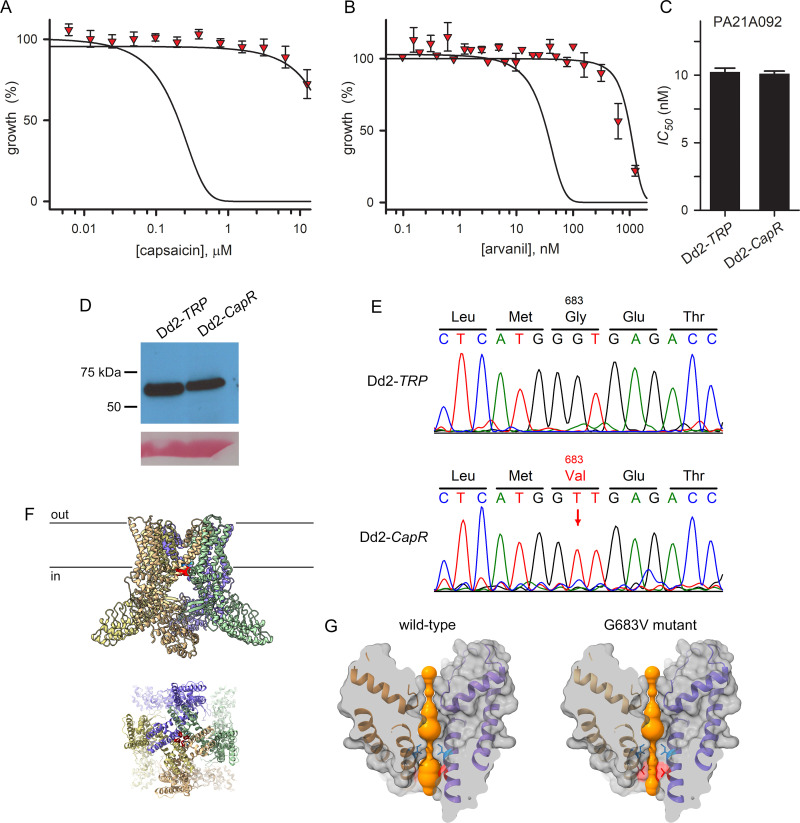
The intracellular human malaria parasite, Plasmodium falciparum, uses the PfATP4 cation pump to maintain Na+ and H+ homeostasis in parasite cytosol. PfATP4 is the target of advanced antimalarial leads, which produce many poorly understood metabolic disturbances within infected erythrocytes. Here, we expressed the mammalian ligand-gated TRPV1 ion channel at the parasite plasma membrane to study ion regulation and examine the effects of cation leak. TRPV1 expression was well-tolerated, consistent with negligible ion flux through the nonactivated channel. TRPV1 ligands produced rapid parasite death in the transfectant line at their activating concentrations, but were harmless to the wild-type parent. Activation triggered cholesterol redistribution at the parasite plasma membrane, reproducing effects of PfATP4 inhibitors and directly implicating cation dysregulation in this process. In contrast to predictions, TRPV1 activation in low Na+ media accentuated parasite killing but a PfATP4 inhibitor had unchanged efficacy. Selection of a ligand-resistant mutant revealed a previously uncharacterized G683V mutation in TRPV1 that occludes the lower channel gate, implicating reduced permeability as a mechanism for parasite resistance to antimalarials targeting ion homeostasis. Our findings provide key insights into malaria parasite ion regulation and will guide mechanism-of-action studies for advanced antimalarial leads that act at the host-pathogen interface.
Copyright: This is an open access article, free of all copyright, and may be freely reproduced, distributed, transmitted, modified, built upon, or otherwise used by anyone for any lawful purpose. The work is made available under the Creative Commons CC0 public domain dedication.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Antimalarials Targeting the Malaria Parasite Cation ATPase P. falciparum ATP4 (PfATP4).Curr Top Med Chem. 2023;23(3):214-226. doi: 10.2174/1568026623666221121154354. Curr Top Med Chem. 2023. PMID: 36411573 Review.
-
Na(+) regulation in the malaria parasite Plasmodium falciparum involves the cation ATPase PfATP4 and is a target of the spiroindolone antimalarials.Cell Host Microbe. 2013 Feb 13;13(2):227-37. doi: 10.1016/j.chom.2012.12.006. Cell Host Microbe. 2013. PMID: 23414762 Free PMC article.
-
Biochemical characterization and chemical inhibition of PfATP4-associated Na+-ATPase activity in Plasmodium falciparum membranes.J Biol Chem. 2018 Aug 24;293(34):13327-13337. doi: 10.1074/jbc.RA118.003640. Epub 2018 Jul 9. J Biol Chem. 2018. PMID: 29986883 Free PMC article.
-
PfATP4 inhibitors in the Medicines for Malaria Venture Malaria Box and Pathogen Box block the schizont-to-ring transition by inhibiting egress rather than invasion.Front Cell Infect Microbiol. 2022 Nov 30;12:1060202. doi: 10.3389/fcimb.2022.1060202. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36530423 Free PMC article.
-
The malaria parasite cation ATPase PfATP4 and its role in the mechanism of action of a new arsenal of antimalarial drugs.Int J Parasitol Drugs Drug Resist. 2015 Aug 27;5(3):149-62. doi: 10.1016/j.ijpddr.2015.07.001. eCollection 2015 Dec. Int J Parasitol Drugs Drug Resist. 2015. PMID: 26401486 Free PMC article. Review.
References
-
- WHO. World malaria report 20212021 9/13/2022.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
