

The unfolded protein response transcription factor XBP1s ameliorates Alzheimer's disease by improving synaptic function and proteostasis
- PMID: 37016577
- PMCID: PMC10362463
- DOI: 10.1016/j.ymthe.2023.03.028
The unfolded protein response transcription factor XBP1s ameliorates Alzheimer's disease by improving synaptic function and proteostasis
Abstract
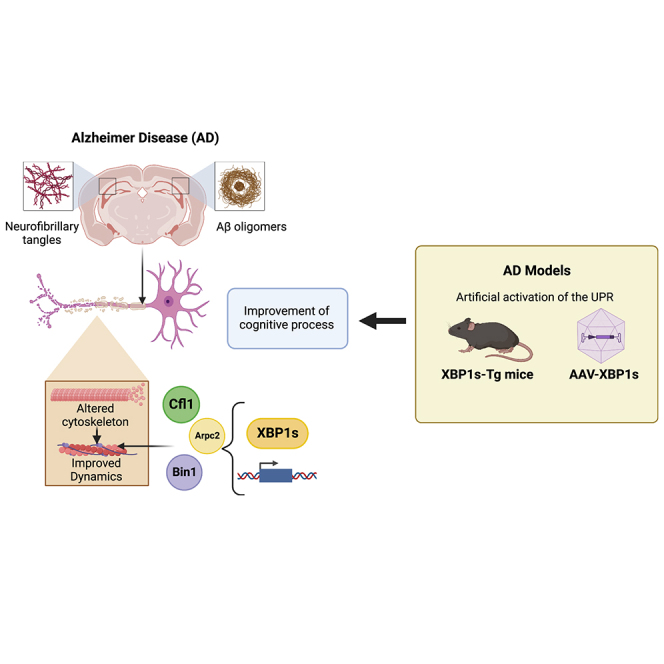
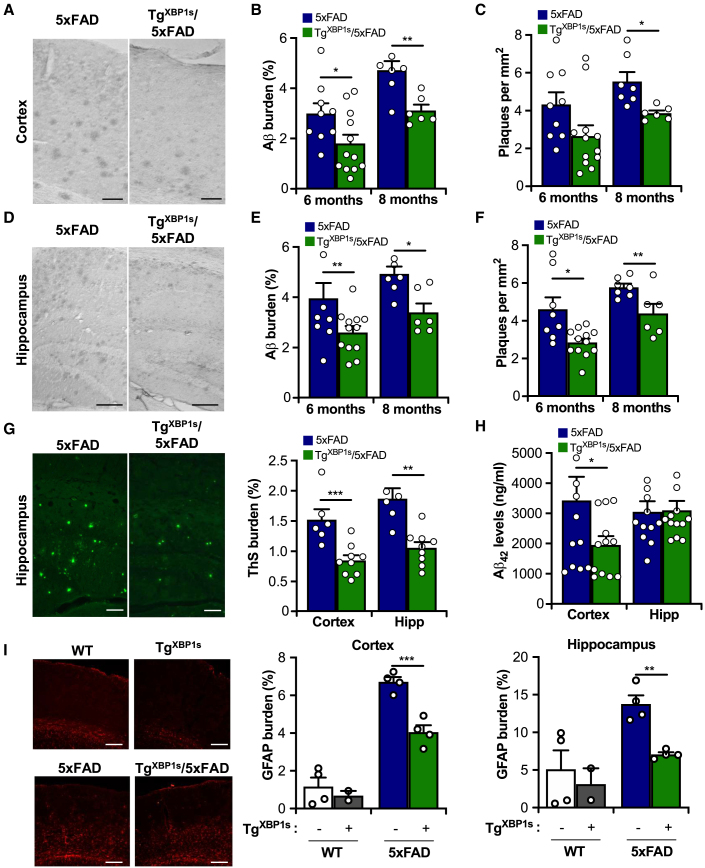
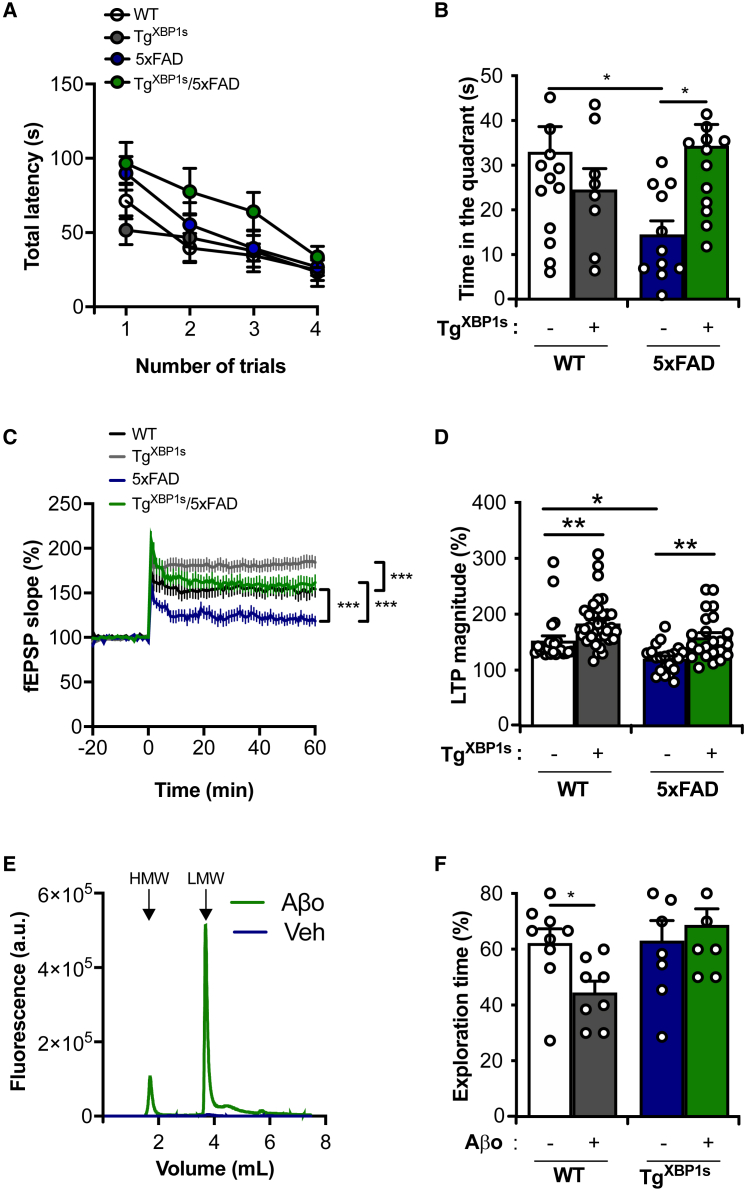
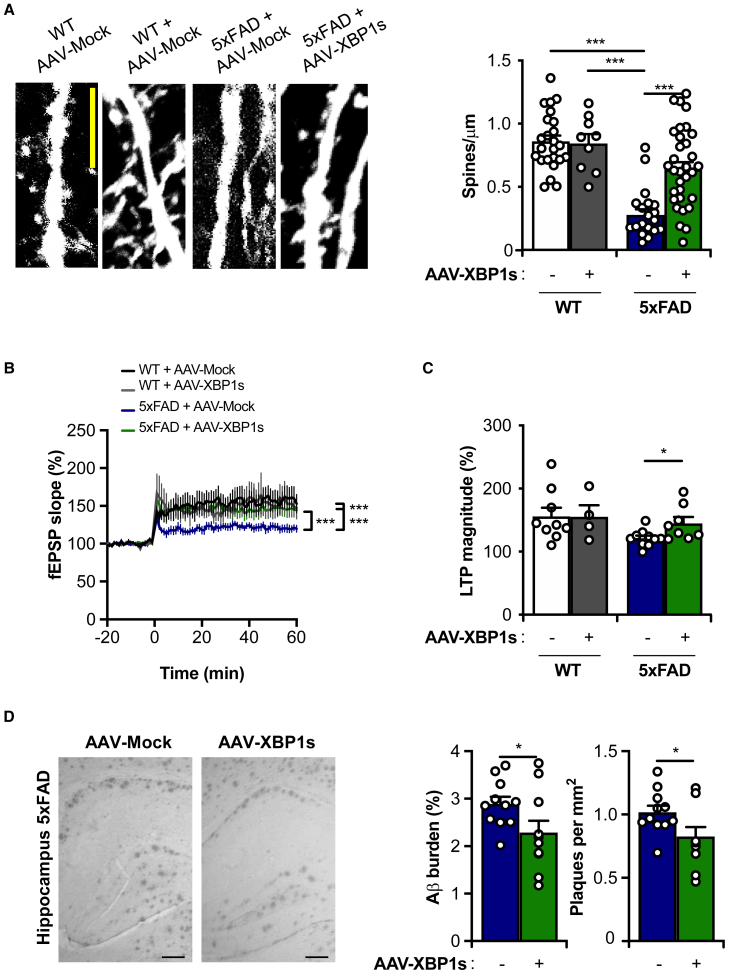
Alteration in the buffering capacity of the proteostasis network is an emerging feature of Alzheimer's disease (AD), highlighting the occurrence of endoplasmic reticulum (ER) stress. The unfolded protein response (UPR) is the main adaptive pathway to cope with protein folding stress at the ER. Inositol-requiring enzyme-1 (IRE1) operates as a central ER stress sensor, enabling the establishment of adaptive and repair programs through the control of the expression of the transcription factor X-box binding protein 1 (XBP1). To artificially enforce the adaptive capacity of the UPR in the AD brain, we developed strategies to express the active form of XBP1 in the brain. Overexpression of XBP1 in the nervous system using transgenic mice reduced the load of amyloid deposits and preserved synaptic and cognitive function. Moreover, local delivery of XBP1 into the hippocampus of an 5xFAD mice using adeno-associated vectors improved different AD features. XBP1 expression corrected a large proportion of the proteomic alterations observed in the AD model, restoring the levels of several synaptic proteins and factors involved in actin cytoskeleton regulation and axonal growth. Our results illustrate the therapeutic potential of targeting UPR-dependent gene expression programs as a strategy to ameliorate AD features and sustain synaptic function.
Keywords: Alzheimer’s disease; ER stress; UPR; XBP1; amyloid β.
Copyright © 2023. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare that no conflicts of interest exist.
Figures


Comment in
-
Beyond the unfolded protein response: Disclosing the role of XBP1s in Alzheimer's disease.Mol Ther. 2023 Jul 5;31(7):1868-1869. doi: 10.1016/j.ymthe.2023.04.015. Epub 2023 May 11. Mol Ther. 2023. PMID: 37172589 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
