

mRNA recognition and packaging by the human transcription-export complex
- PMID: 37020021
- PMCID: PMC7614608
- DOI: 10.1038/s41586-023-05904-0
mRNA recognition and packaging by the human transcription-export complex
Abstract
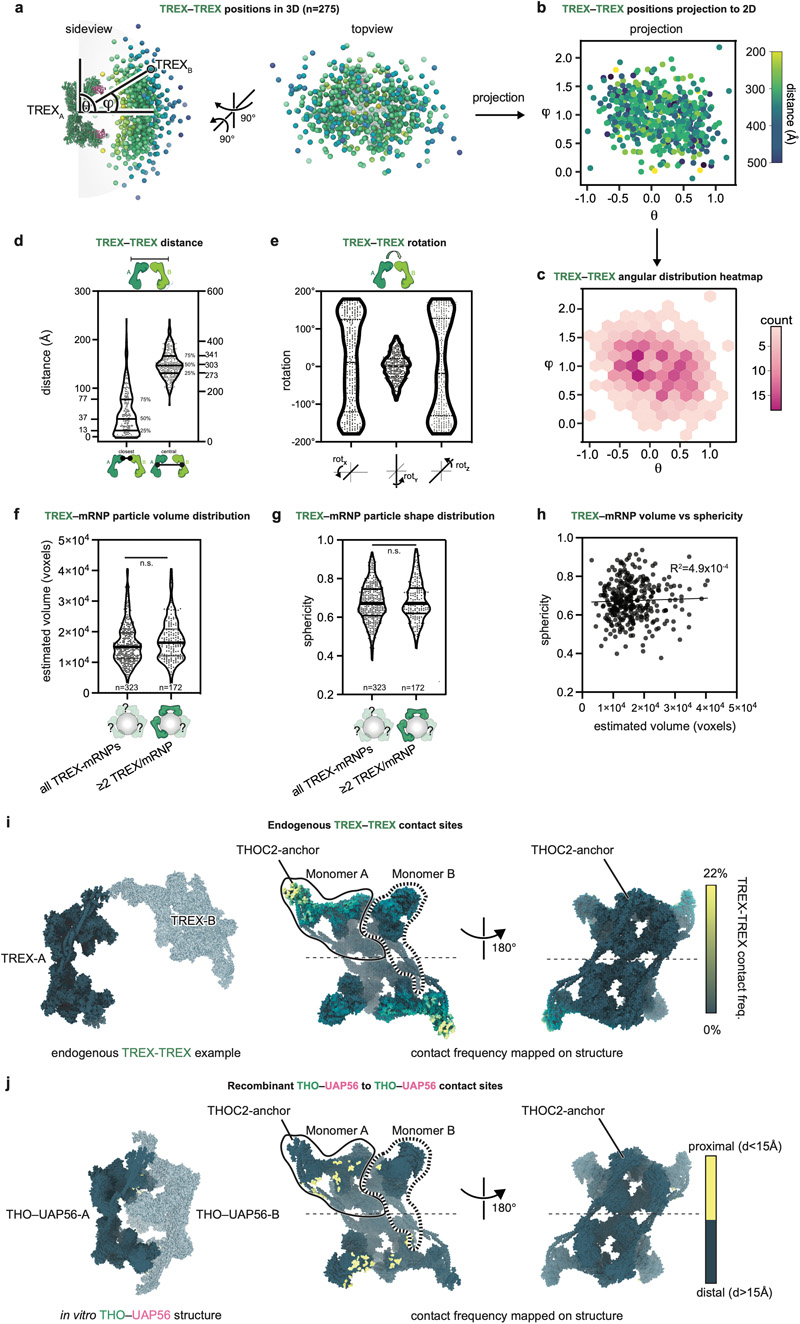
Newly made mRNAs are processed and packaged into mature ribonucleoprotein complexes (mRNPs) and are recognized by the essential transcription-export complex (TREX) for nuclear export1,2. However, the mechanisms of mRNP recognition and three-dimensional mRNP organization are poorly understood3. Here we report cryo-electron microscopy and tomography structures of reconstituted and endogenous human mRNPs bound to the 2-MDa TREX complex. We show that mRNPs are recognized through multivalent interactions between the TREX subunit ALYREF and mRNP-bound exon junction complexes. Exon junction complexes can multimerize through ALYREF, which suggests a mechanism for mRNP organization. Endogenous mRNPs form compact globules that are coated by multiple TREX complexes. These results reveal how TREX may simultaneously recognize, compact and protect mRNAs to promote their packaging for nuclear export. The organization of mRNP globules provides a framework to understand how mRNP architecture facilitates mRNA biogenesis and export.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures














Similar articles
-
Structure of the human core transcription-export complex reveals a hub for multivalent interactions.Elife. 2020 Nov 16;9:e61503. doi: 10.7554/eLife.61503. Elife. 2020. PMID: 33191911 Free PMC article.
-
Structures and mRNP remodeling mechanism of the TREX-2 complex.Structure. 2025 Mar 6;33(3):566-582.e6. doi: 10.1016/j.str.2024.12.019. Epub 2025 Jan 24. Structure. 2025. PMID: 39862860
-
Structural mechanism of DDX39B regulation by human TREX-2 and a related complex in mRNP remodeling.Nat Commun. 2025 Jul 1;16(1):5471. doi: 10.1038/s41467-025-60547-1. Nat Commun. 2025. PMID: 40595470 Free PMC article.
-
Early formation of mRNP: license for export or quality control?Mol Cell. 2003 May;11(5):1129-38. doi: 10.1016/s1097-2765(03)00191-6. Mol Cell. 2003. PMID: 12769839 Review.
-
mRNA accessibility within mRNPs as a determinant of gene expression.Trends Biochem Sci. 2024 Mar;49(3):199-207. doi: 10.1016/j.tibs.2023.11.003. Epub 2023 Dec 8. Trends Biochem Sci. 2024. PMID: 38071089 Free PMC article. Review.
Cited by
-
Predicting the Effect of Single Mutations on Protein Stability and Binding with Respect to Types of Mutations.Int J Mol Sci. 2023 Jul 28;24(15):12073. doi: 10.3390/ijms241512073. Int J Mol Sci. 2023. PMID: 37569449 Free PMC article.
-
Subcytoplasmic location of translation controls protein output.Mol Cell. 2023 Dec 21;83(24):4509-4523.e11. doi: 10.1016/j.molcel.2023.11.025. Mol Cell. 2023. PMID: 38134885 Free PMC article.
-
The molecular architecture of the nuclear basket.Cell. 2024 Sep 19;187(19):5267-5281.e13. doi: 10.1016/j.cell.2024.07.020. Epub 2024 Aug 9. Cell. 2024. PMID: 39127037 Free PMC article.
-
Large-scale map of RNA-binding protein interactomes across the mRNA life cycle.Mol Cell. 2024 Oct 3;84(19):3790-3809.e8. doi: 10.1016/j.molcel.2024.08.030. Epub 2024 Sep 19. Mol Cell. 2024. PMID: 39303721
-
Cryo-EM structure of the CBC-ALYREF complex.Elife. 2024 Sep 16;12:RP91432. doi: 10.7554/eLife.91432. Elife. 2024. PMID: 39282949 Free PMC article.
References
-
- Köhler A, Hurt E. Exporting RNA from the nucleus to the cytoplasm. Nat Rev Mol Cell Biol. 2007;8:761–773. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
