

Over and above frequency: Gamma oscillations as units of neural circuit operations
- PMID: 37023717
- PMCID: PMC7614431
- DOI: 10.1016/j.neuron.2023.02.026
Over and above frequency: Gamma oscillations as units of neural circuit operations
Abstract
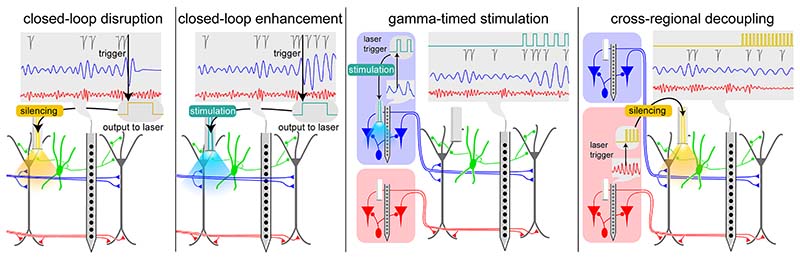
Gamma oscillations (∼30-150 Hz) are widespread correlates of neural circuit functions. These network activity patterns have been described across multiple animal species, brain structures, and behaviors, and are usually identified based on their spectral peak frequency. Yet, despite intensive investigation, whether gamma oscillations implement causal mechanisms of specific brain functions or represent a general dynamic mode of neural circuit operation remains unclear. In this perspective, we review recent advances in the study of gamma oscillations toward a deeper understanding of their cellular mechanisms, neural pathways, and functional roles. We discuss that a given gamma rhythm does not per se implement any specific cognitive function but rather constitutes an activity motif reporting the cellular substrates, communication channels, and computational operations underlying information processing in its generating brain circuit. Accordingly, we propose shifting the attention from a frequency-based to a circuit-level definition of gamma oscillations.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
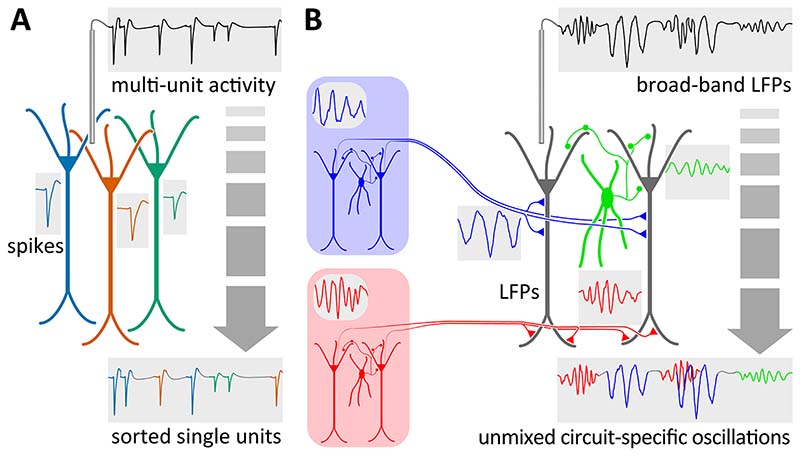
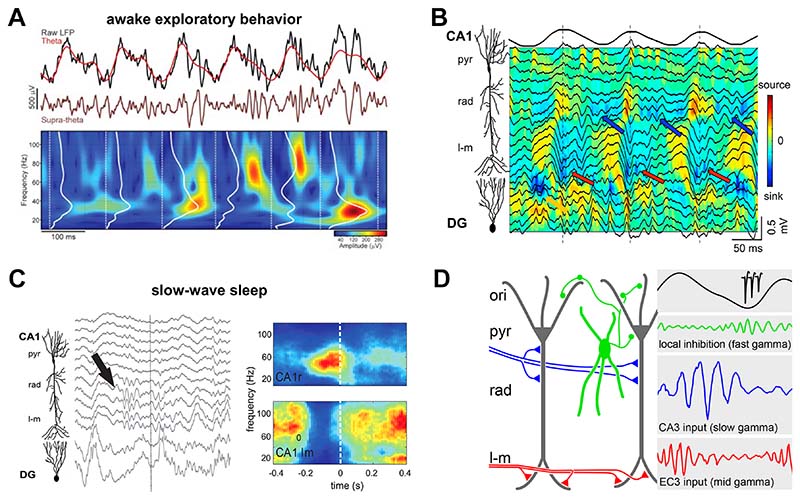
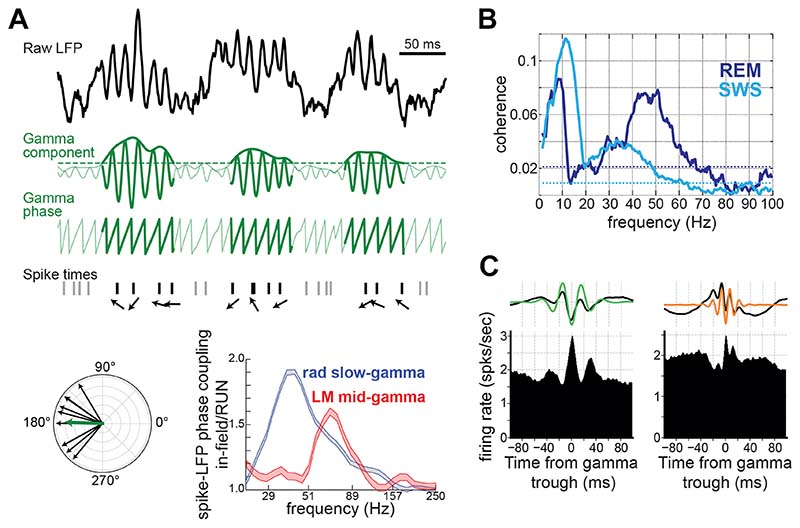
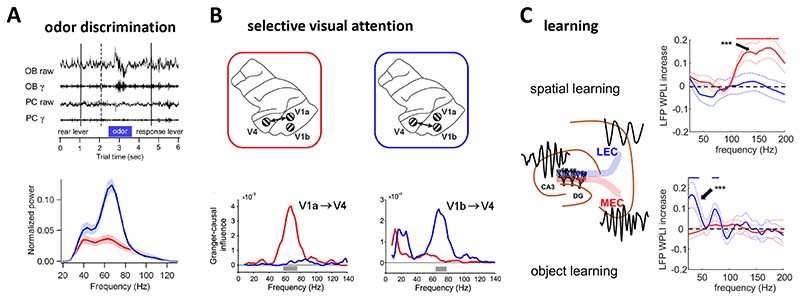
Similar articles
-
Snapshots of the Brain in Action: Local Circuit Operations through the Lens of γ Oscillations.J Neurosci. 2016 Oct 12;36(41):10496-10504. doi: 10.1523/JNEUROSCI.1021-16.2016. J Neurosci. 2016. PMID: 27733601 Free PMC article.
-
Functions of gamma-band synchronization in cognition: from single circuits to functional diversity across cortical and subcortical systems.Eur J Neurosci. 2014 Jun;39(11):1982-99. doi: 10.1111/ejn.12606. Epub 2014 May 8. Eur J Neurosci. 2014. PMID: 24809619 Review.
-
Gamma and Beta Oscillations Define a Sequence of Neurocognitive Modes Present in Odor Processing.J Neurosci. 2016 Jul 20;36(29):7750-67. doi: 10.1523/JNEUROSCI.0569-16.2016. J Neurosci. 2016. PMID: 27445151 Free PMC article.
-
Cross-frequency coupling between gamma oscillations and deep brain stimulation frequency in Parkinson's disease.Brain. 2020 Dec 5;143(11):3393-3407. doi: 10.1093/brain/awaa297. Brain. 2020. PMID: 33150359 Free PMC article.
-
Deciphering the code: Identifying true gamma neural oscillations.Exp Neurol. 2022 Nov;357:114205. doi: 10.1016/j.expneurol.2022.114205. Epub 2022 Aug 17. Exp Neurol. 2022. PMID: 35985554 Review.
Cited by
-
Transcranial alternating current stimulation in affecting cognitive impairment in psychiatric disorders: a review.Eur Arch Psychiatry Clin Neurosci. 2024 Jun;274(4):803-826. doi: 10.1007/s00406-023-01687-7. Epub 2023 Sep 8. Eur Arch Psychiatry Clin Neurosci. 2024. PMID: 37682331 Free PMC article. Review.
-
Fast, accurate, and interpretable decoding of electrocorticographic signals using dynamic mode decomposition.Commun Biol. 2024 May 18;7(1):595. doi: 10.1038/s42003-024-06294-3. Commun Biol. 2024. PMID: 38762683 Free PMC article.
-
Exploring a Novel Hypothesis: Could the Eye Function as a Radar or Ultrasound Device in Depth and Distance Perception? Neurophysiological Insights.Life (Basel). 2025 Mar 25;15(4):536. doi: 10.3390/life15040536. Life (Basel). 2025. PMID: 40283091 Free PMC article.
-
Modulation of stress-, pain-, and alcohol-related behaviors by perineuronal nets.Neurobiol Stress. 2024 Nov 14;33:100692. doi: 10.1016/j.ynstr.2024.100692. eCollection 2024 Nov. Neurobiol Stress. 2024. PMID: 39691634 Free PMC article.
-
Large-scale coupling of prefrontal activity patterns as a mechanism for cognitive control in health and disease: evidence from rodent models.Front Neural Circuits. 2024 Apr 4;18:1286111. doi: 10.3389/fncir.2024.1286111. eCollection 2024. Front Neural Circuits. 2024. PMID: 38638163 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
