

Intercellular exchange of Wnt ligands reduces cell population heterogeneity during embryogenesis
- PMID: 37024462
- PMCID: PMC10079677
- DOI: 10.1038/s41467-023-37350-x
Intercellular exchange of Wnt ligands reduces cell population heterogeneity during embryogenesis
Abstract
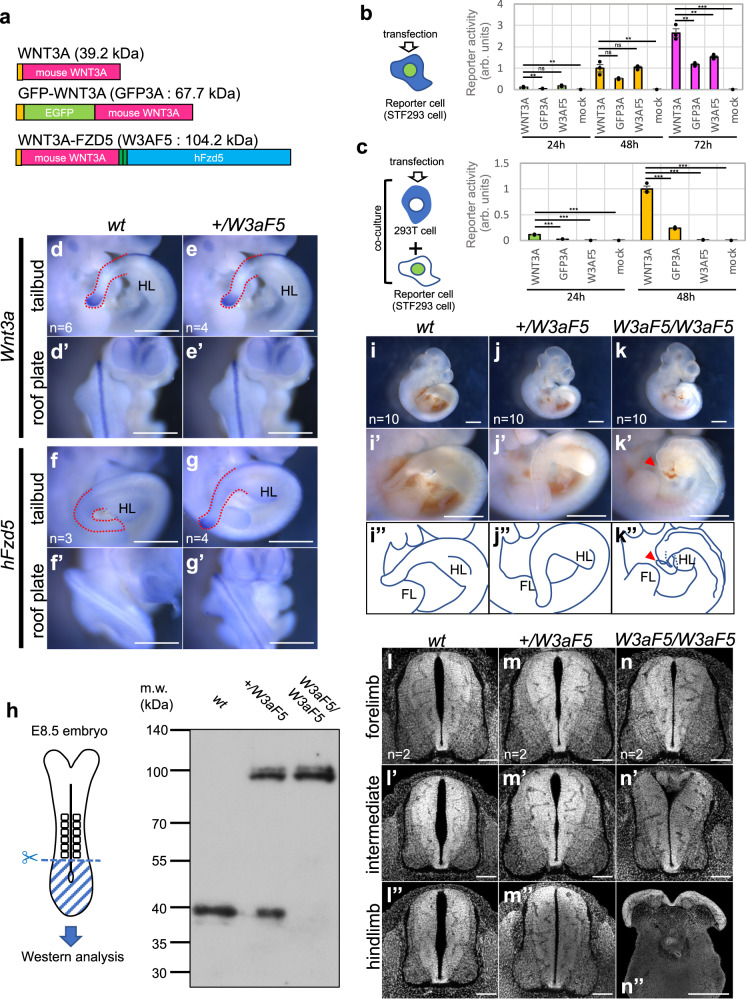
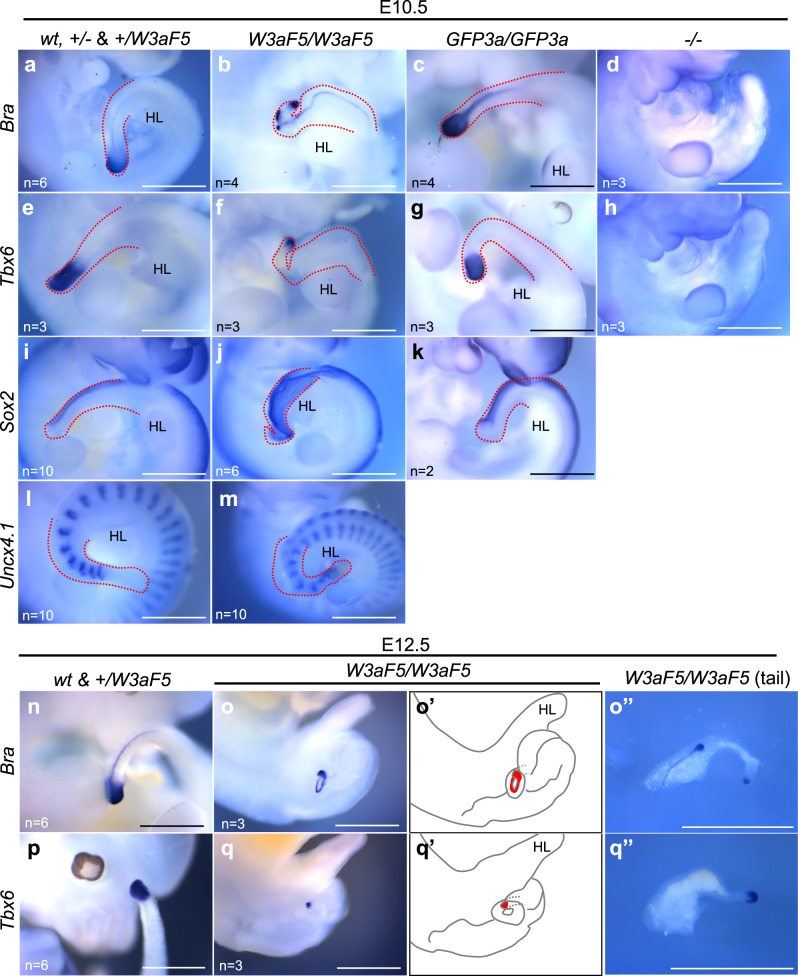
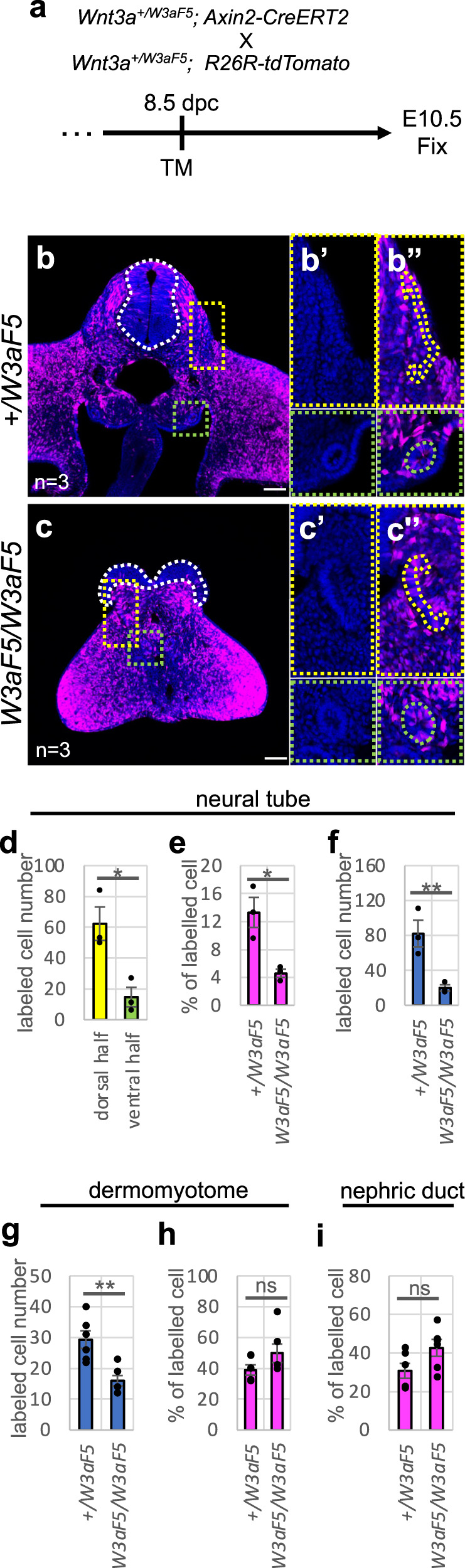
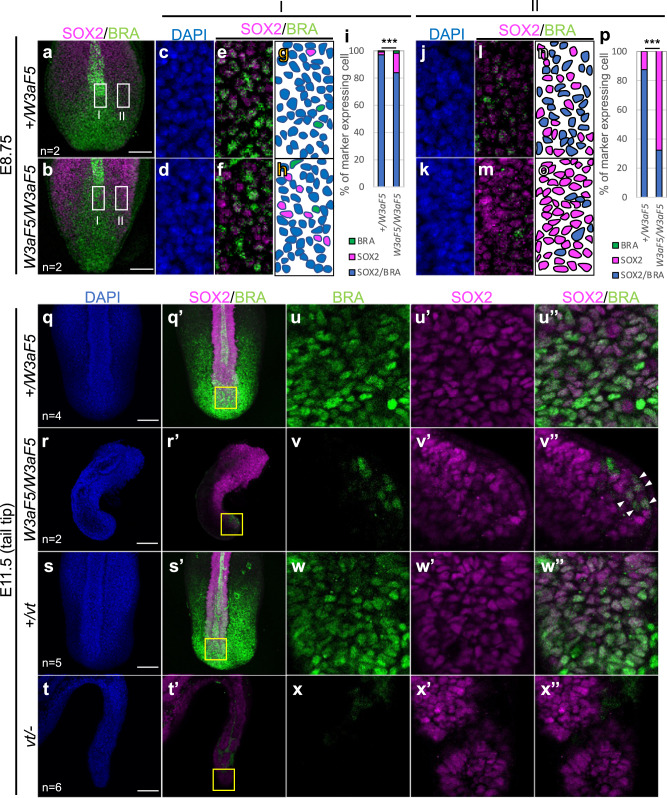
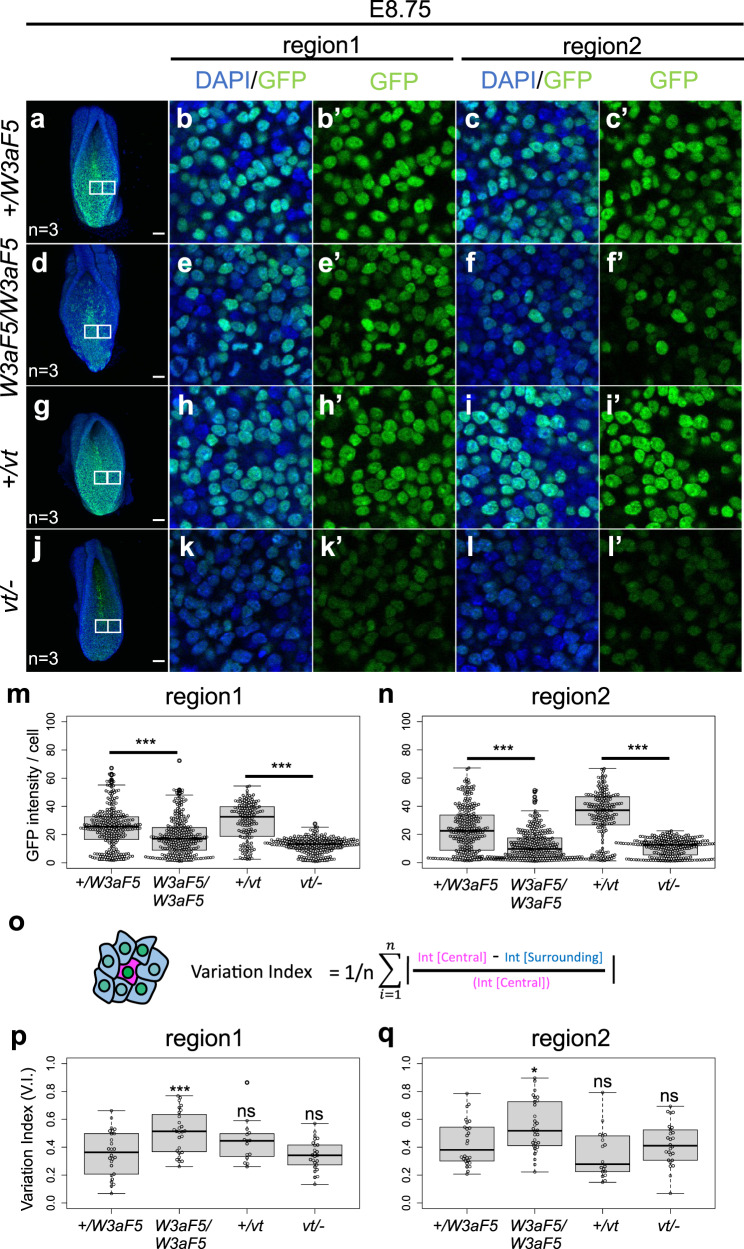
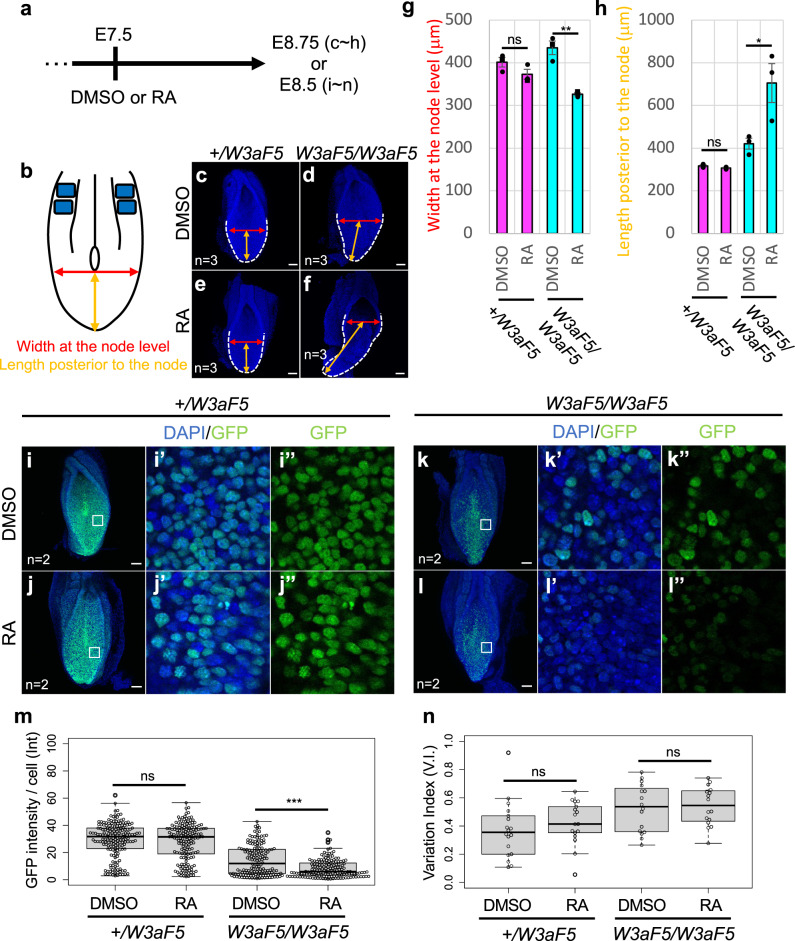
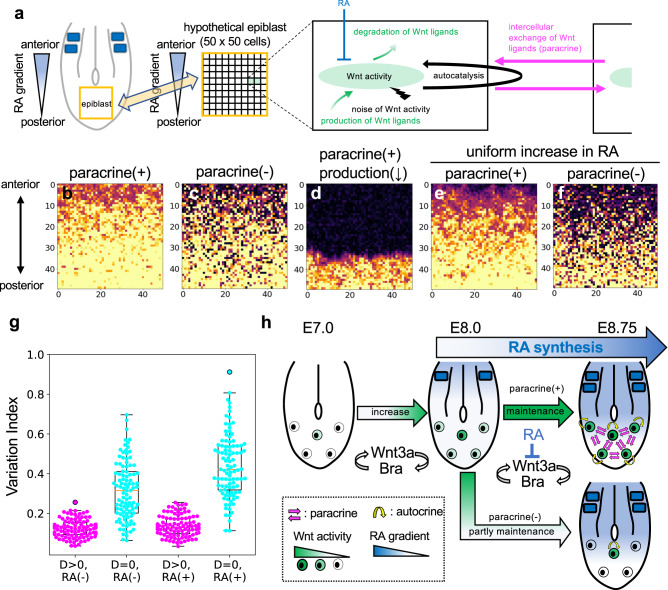
Wnt signaling is required to maintain bipotent progenitors for neural and paraxial mesoderm cells, the neuromesodermal progenitor (NMP) cells that reside in the epiblast and tailbud. Since epiblast/tailbud cells receive Wnt ligands produced by one another, this exchange may average out the heterogeneity of Wnt signaling levels among these cells. Here, we examined this possibility by replacing endogenous Wnt3a with a receptor-fused form that activates signaling in producing cells, but not in neighboring cells. Mutant mouse embryos show a unique phenotype in which maintenance of many NMP cells is impaired, although some cells persist for long periods. The epiblast cell population of these embryos increases heterogeneity in Wnt signaling levels as embryogenesis progresses and are sensitive to retinoic acid, an endogenous antagonist of NMP maintenance. Thus, mutual intercellular exchange of Wnt ligands in the epiblast cell population reduces heterogeneity and achieves robustness to environmental stress.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
The Chick Caudolateral Epiblast Acts as a Permissive Niche for Generating Neuromesodermal Progenitor Behaviours.Cells Tissues Organs. 2018;205(5-6):320-330. doi: 10.1159/000494769. Epub 2018 Dec 5. Cells Tissues Organs. 2018. PMID: 30517924 Free PMC article.
-
Wnt/β-catenin and FGF signalling direct the specification and maintenance of a neuromesodermal axial progenitor in ensembles of mouse embryonic stem cells.Development. 2014 Nov;141(22):4243-53. doi: 10.1242/dev.112979. Development. 2014. PMID: 25371361 Free PMC article.
-
Inverse agonism of retinoic acid receptors directs epiblast cells into the paraxial mesoderm lineage.Stem Cell Res. 2018 Jul;30:85-95. doi: 10.1016/j.scr.2018.05.016. Epub 2018 May 22. Stem Cell Res. 2018. PMID: 29807258 Free PMC article.
-
The Origin and Regulation of Neuromesodermal Progenitors (NMPs) in Embryos.Cells. 2024 Mar 21;13(6):549. doi: 10.3390/cells13060549. Cells. 2024. PMID: 38534393 Free PMC article. Review.
-
Wnt/ß-catenin signalling and the dynamics of fate decisions in early mouse embryos and embryonic stem (ES) cells.Semin Cell Dev Biol. 2015 Dec;47-48:101-9. doi: 10.1016/j.semcdb.2015.08.011. Epub 2015 Aug 29. Semin Cell Dev Biol. 2015. PMID: 26321498 Free PMC article. Review.
Cited by
-
The Clock and Wavefront Self-Organizing model recreates the dynamics of mouse somitogenesis in vivo and in vitro.Development. 2024 May 15;151(10):dev202606. doi: 10.1242/dev.202606. Epub 2024 May 16. Development. 2024. PMID: 38742434 Free PMC article.
-
Wnt-dependent mechanism of the apical constriction of roof plate cells in developing mouse spinal cord.Front Cell Dev Biol. 2025 Jul 7;13:1571770. doi: 10.3389/fcell.2025.1571770. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40692752 Free PMC article.
-
Rho-Associated Protein Kinase Activity Is Required for Tissue Homeostasis in the Xenopus laevis Ciliated Epithelium.J Dev Biol. 2024 Jun 11;12(2):17. doi: 10.3390/jdb12020017. J Dev Biol. 2024. PMID: 38921484 Free PMC article.
-
The role of the striatin family proteins in hippo signaling and cellular regulation.Cell Biosci. 2025 Aug 19;15(1):119. doi: 10.1186/s13578-025-01461-3. Cell Biosci. 2025. PMID: 40830981 Free PMC article.
-
Understanding and manipulating extracellular behaviors of Wnt ligands.In Vitro Cell Dev Biol Anim. 2024 May;60(5):441-448. doi: 10.1007/s11626-024-00856-9. Epub 2024 Feb 20. In Vitro Cell Dev Biol Anim. 2024. PMID: 38379096 Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
