

Multilevel interrogation of H3.3 reveals a primordial role in transcription regulation
- PMID: 37024975
- PMCID: PMC10080907
- DOI: 10.1186/s13072-023-00484-9
Multilevel interrogation of H3.3 reveals a primordial role in transcription regulation
Abstract
Background: Eukaryotic cells can rapidly adjust their transcriptional profile in response to molecular needs. Such dynamic regulation is, in part, achieved through epigenetic modifications and selective incorporation of histone variants into chromatin. H3.3 is the ancestral H3 variant with key roles in regulating chromatin states and transcription. Although H3.3 has been well studied in metazoans, information regarding the assembly of H3.3 onto chromatin and its possible role in transcription regulation remain poorly documented outside of Opisthokonts.
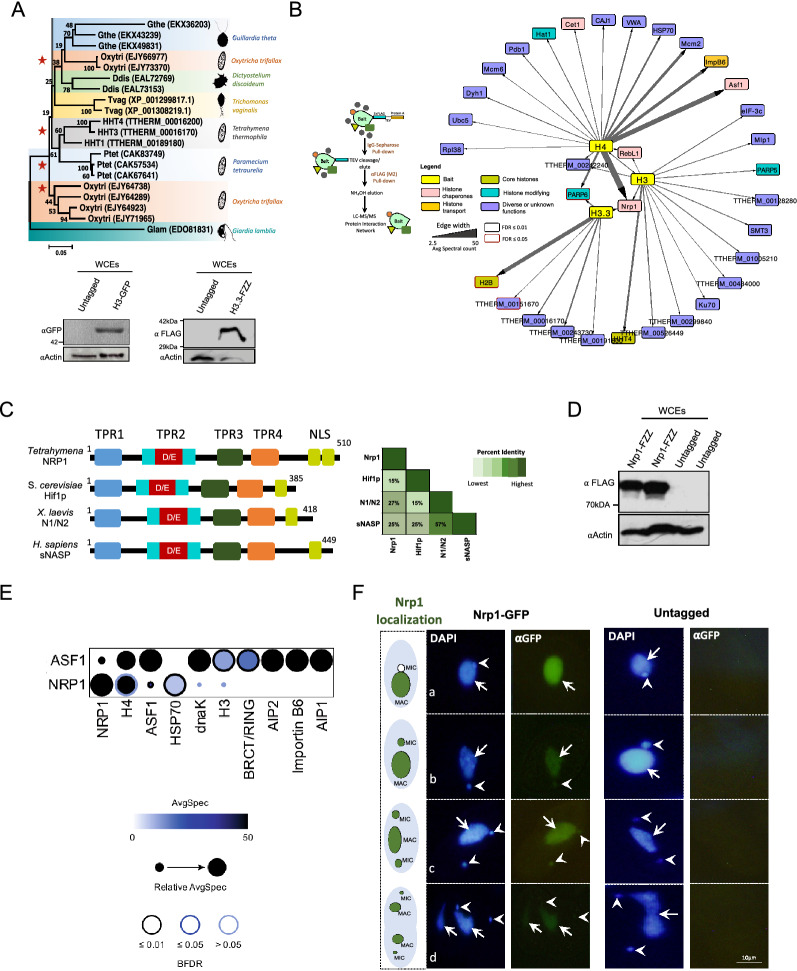
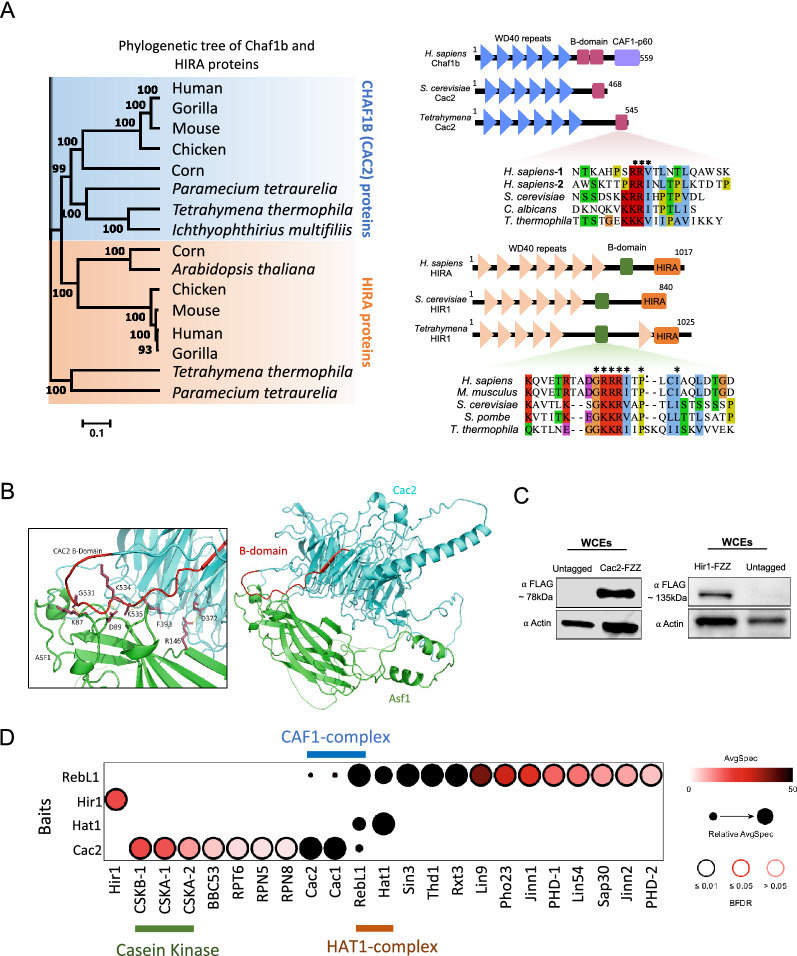
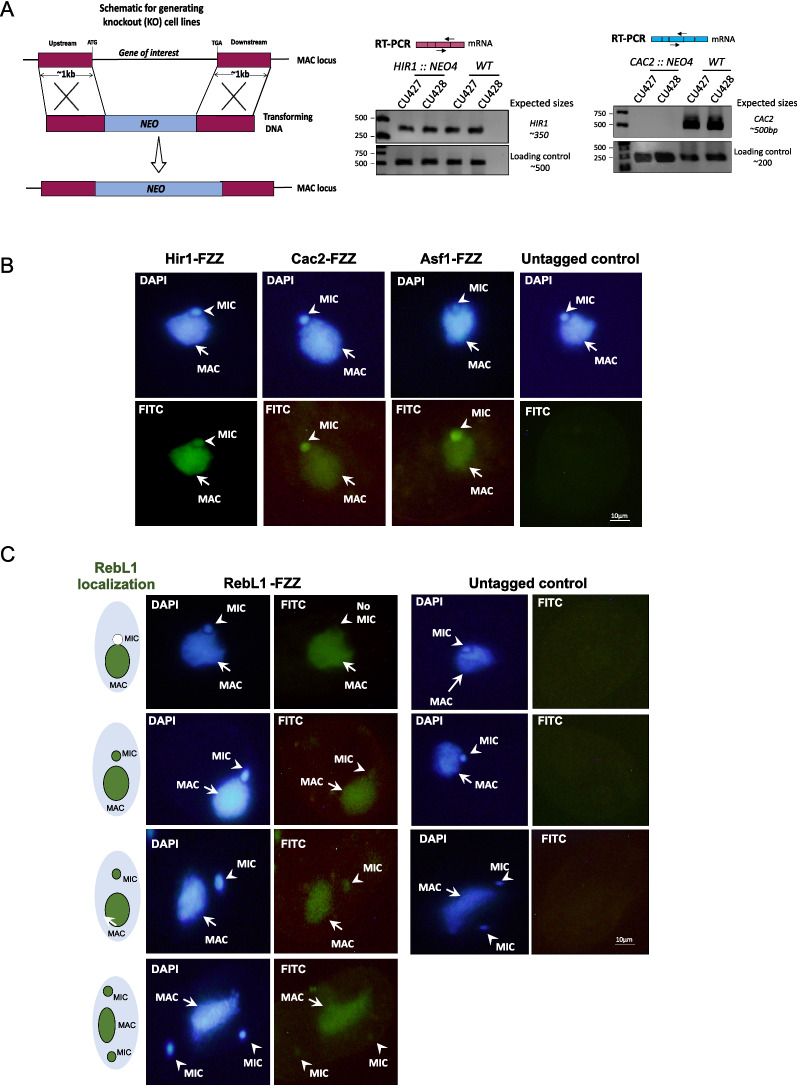
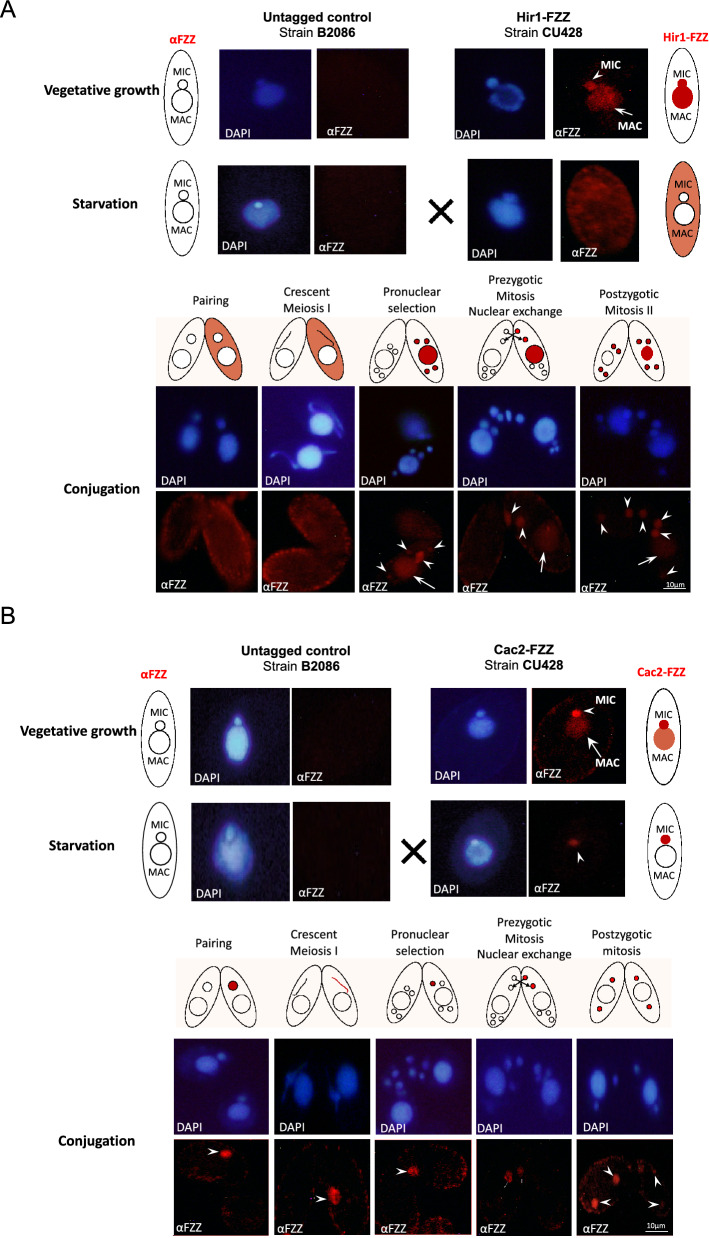
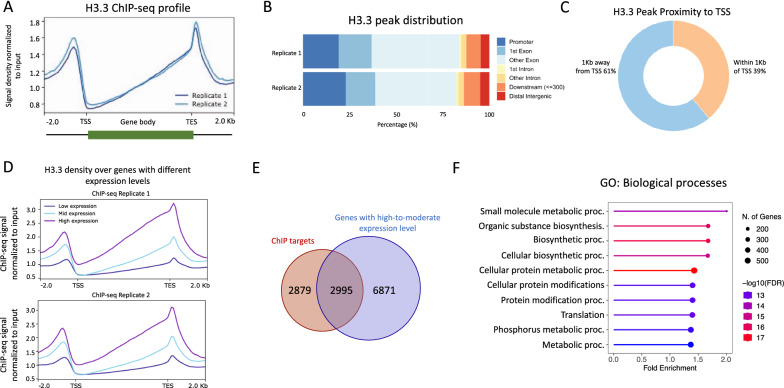
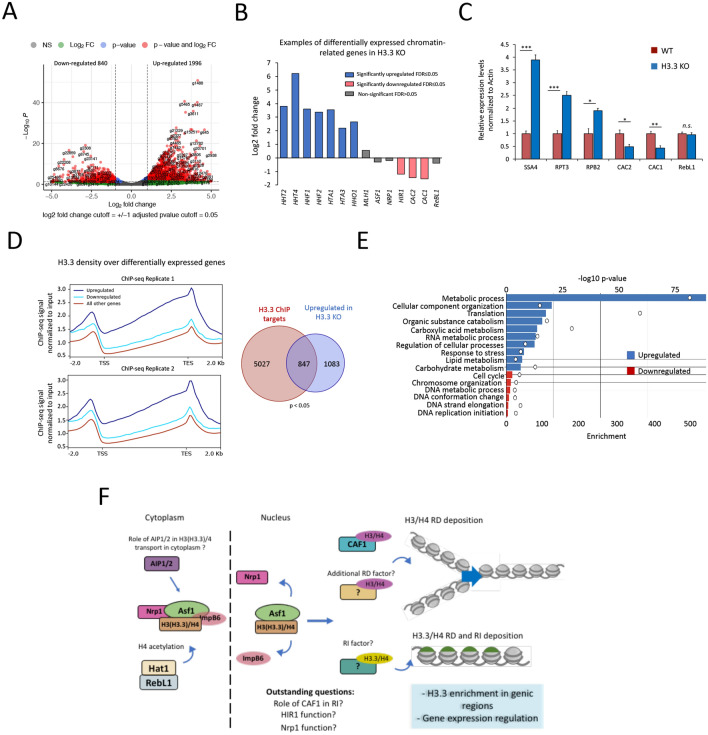
Results: We used the nuclear dimorphic ciliate protozoan, Tetrahymena thermophila, to investigate the dynamics of H3 variant function in evolutionarily divergent eukaryotes. Functional proteomics and immunofluorescence analyses of H3.1 and H3.3 revealed a highly conserved role for Nrp1 and Asf1 histone chaperones in nuclear influx of histones. Cac2, a putative subunit of H3.1 deposition complex CAF1, is not required for growth, whereas the expression of the putative ortholog of the H3.3-specific chaperone Hir1 is essential in Tetrahymena. Our results indicate that Cac2 and Hir1 have distinct localization patterns during different stages of the Tetrahymena life cycle and suggest that Cac2 might be dispensable for chromatin assembly. ChIP-seq experiments in growing Tetrahymena show H3.3 enrichment over the promoters, gene bodies, and transcription termination sites of highly transcribed genes. H3.3 knockout followed by RNA-seq reveals large-scale transcriptional alterations in functionally important genes.
Conclusion: Our results provide an evolutionary perspective on H3.3's conserved role in maintaining the transcriptional landscape of cells and on the emergence of specialized chromatin assembly pathways.
Keywords: Asf1; CAF1; Chromatin; Epigenetics; Functional proteomics; H3.3; HIRA; Histone variant; NASP; RBBP4/7; Tetrahymena.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Conserved Asf1-importin β physical interaction in growth and sexual development in the ciliate Tetrahymena thermophila.J Proteomics. 2013 Dec 6;94:311-26. doi: 10.1016/j.jprot.2013.09.018. Epub 2013 Oct 11. J Proteomics. 2013. PMID: 24120531
-
Histone chaperone ASF1 mediates H3.3-H4 deposition in Arabidopsis.Nat Commun. 2022 Nov 15;13(1):6970. doi: 10.1038/s41467-022-34648-0. Nat Commun. 2022. PMID: 36379930 Free PMC article.
-
Proteomic Analysis of Histones H2A/H2B and Variant Hv1 in Tetrahymena thermophila Reveals an Ancient Network of Chaperones.Mol Biol Evol. 2019 May 1;36(5):1037-1055. doi: 10.1093/molbev/msz039. Mol Biol Evol. 2019. PMID: 30796450 Free PMC article.
-
A Molecular Prospective for HIRA Complex Assembly and H3.3-Specific Histone Chaperone Function.J Mol Biol. 2017 Jun 30;429(13):1924-1933. doi: 10.1016/j.jmb.2016.11.010. Epub 2016 Nov 19. J Mol Biol. 2017. PMID: 27871933 Free PMC article. Review.
-
Dynamics of histone variant H3.3 and its coregulation with H2A.Z at enhancers and promoters.Nucleus. 2014 Jan-Feb;5(1):21-7. doi: 10.4161/nucl.28067. Epub 2014 Feb 3. Nucleus. 2014. PMID: 24637397 Free PMC article. Review.
Cited by
-
Bromodomain proteins IBD1 and IBD2 link histone acetylation to SWR1- and INO80-mediated H2A.Z regulation in Tetrahymena.Epigenetics Chromatin. 2025 Aug 6;18(1):51. doi: 10.1186/s13072-025-00614-5. Epigenetics Chromatin. 2025. PMID: 40764940 Free PMC article.
-
Independent and Complementary Functions of Caf1b and Hir1 for Chromatin Assembly in Tetrahymena thermophila.Cells. 2023 Dec 13;12(24):2828. doi: 10.3390/cells12242828. Cells. 2023. PMID: 38132148 Free PMC article.
-
RebL1 is required for macronuclear structure stability and gametogenesis in Tetrahymena thermophila.Mar Life Sci Technol. 2024 Mar 26;6(2):183-197. doi: 10.1007/s42995-024-00219-z. eCollection 2024 May. Mar Life Sci Technol. 2024. PMID: 38827131 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
