

Comparative Effects of Domain-Specific Human Monoclonal Antibodies Against LGI1 on Neuronal Excitability
- PMID: 37028941
- PMCID: PMC10099296
- DOI: 10.1212/NXI.0000000000200096
Comparative Effects of Domain-Specific Human Monoclonal Antibodies Against LGI1 on Neuronal Excitability
Abstract
Background and objectives: Autoantibodies to leucine-rich glioma inactivated protein 1 (LGI1) cause an autoimmune limbic encephalitis with frequent focal seizures and anterograde memory dysfunction. LGI1 is a neuronal secreted linker protein with 2 functional domains: the leucine-rich repeat (LRR) and epitempin (EPTP) regions. LGI1 autoantibodies are known to interfere with presynaptic function and neuronal excitability; however, their epitope-specific mechanisms are incompletely understood.
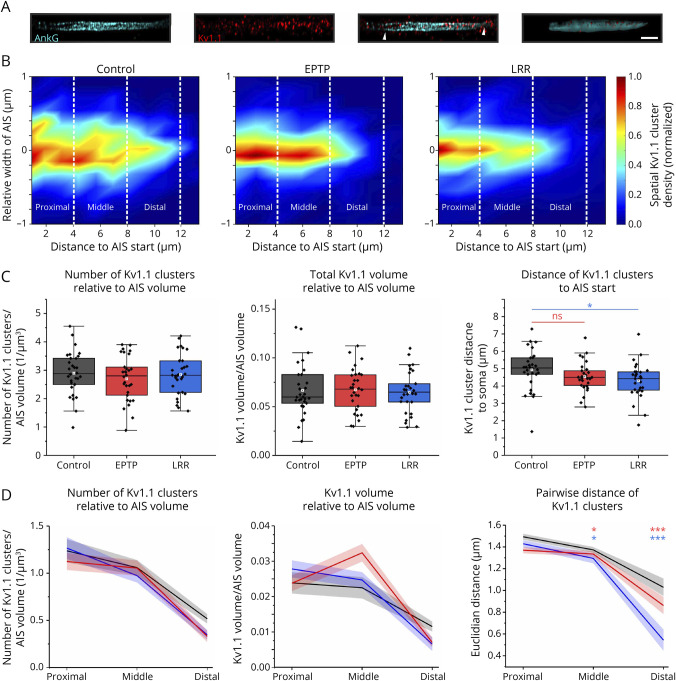
Methods: We used patient-derived monoclonal autoantibodies (mAbs), which target either LRR or EPTP domains of LGI1 to investigate long-term antibody-induced alteration of neuronal function. LRR- and EPTP-specific effects were evaluated by patch-clamp recordings in cultured hippocampal neurons and compared with biophysical neuron modeling. Kv1.1 channel clustering at the axon initial segment (AIS) was quantified by immunocytochemistry and structured illumination microscopy techniques.
Results: Both EPTP and LRR domain-specific mAbs decreased the latency of first somatic action potential firing. However, only the LRR-specific mAbs increased the number of action potential firing together with enhanced initial instantaneous frequency and promoted spike-frequency adaptation, which were less pronounced after the EPTP mAb. This also led to an effective reduction in the slope of ramp-like depolarization in the subthreshold response, suggesting Kv1 channel dysfunction. A biophysical model of a hippocampal neuron corroborated experimental results and suggests that an isolated reduction of the conductance of Kv1-mediated K+ currents largely accounts for the antibody-induced alterations in the initial firing phase and spike-frequency adaptation. Furthermore, Kv1.1 channel density was spatially redistributed from the distal toward the proximal site of AIS under LRR mAb treatment and, to a lesser extant, under EPTP mAb.
Discussion: These findings indicate an epitope-specific pathophysiology of LGI1 autoantibodies. The pronounced neuronal hyperexcitability and SFA together with dropped slope of ramp-like depolarization after LRR-targeted interference suggest disruption of LGI1-dependent clustering of K+ channel complexes. Moreover, considering the effective triggering of action potentials at the distal AIS, the altered spatial distribution of Kv1.1 channel density may contribute to these effects through impairing neuronal control of action potential initiation and synaptic integration.
Copyright © 2023 The Author(s). Published by Wolters Kluwer Health, Inc. on behalf of the American Academy of Neurology.
Conflict of interest statement
C. Geis received honoraria and/or research support from Alexion and Roche. S.R. Irani is an inventor on Diagnostic Strategy to improve specificity of CASPR2 antibody detection (PCT/G82019/051257) and receives royalties on a licensed patent application for LGI1/CASPR2 testing as coapplicant (PCT/GB2009/051441) entitled “Neurological Autoimmune Disorders”. S.R. Irani has received honoraria and/or research support from UCB, Immunovant, MedImmune, Roche, Janssen, Cerebral therapeutics, CSL Behring, and ONO Pharma. The other authors report no disclosures relevant to the manuscript. Go to
Figures





References
-
- Irani SR, Alexander S, Waters P, et al. . Antibodies to Kv1 potassium channel-complex proteins leucine-rich, glioma inactivated 1 protein and contactin-associated protein-2 in limbic encephalitis, Morvan's syndrome and acquired neuromyotonia. Brain. 2010;133(9):2734-2748. doi. 10.1093/brain/awq213 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials