

METTL14 is a chromatin regulator independent of its RNA N6-methyladenosine methyltransferase activity
- PMID: 37030005
- PMCID: PMC10501186
- DOI: 10.1093/procel/pwad009
METTL14 is a chromatin regulator independent of its RNA N6-methyladenosine methyltransferase activity
Abstract
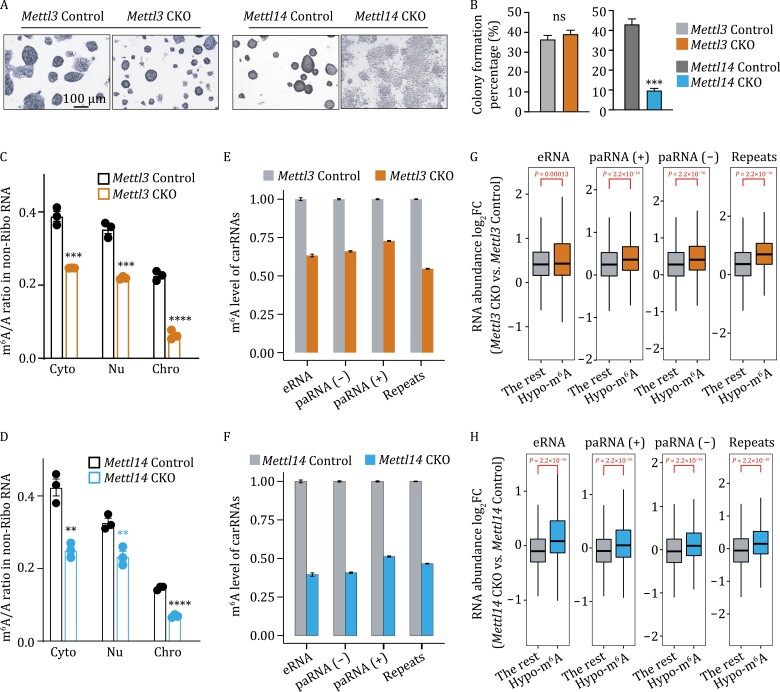
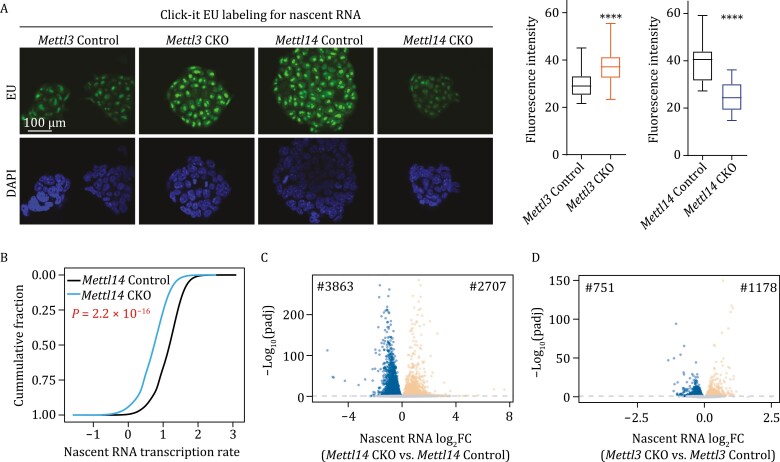
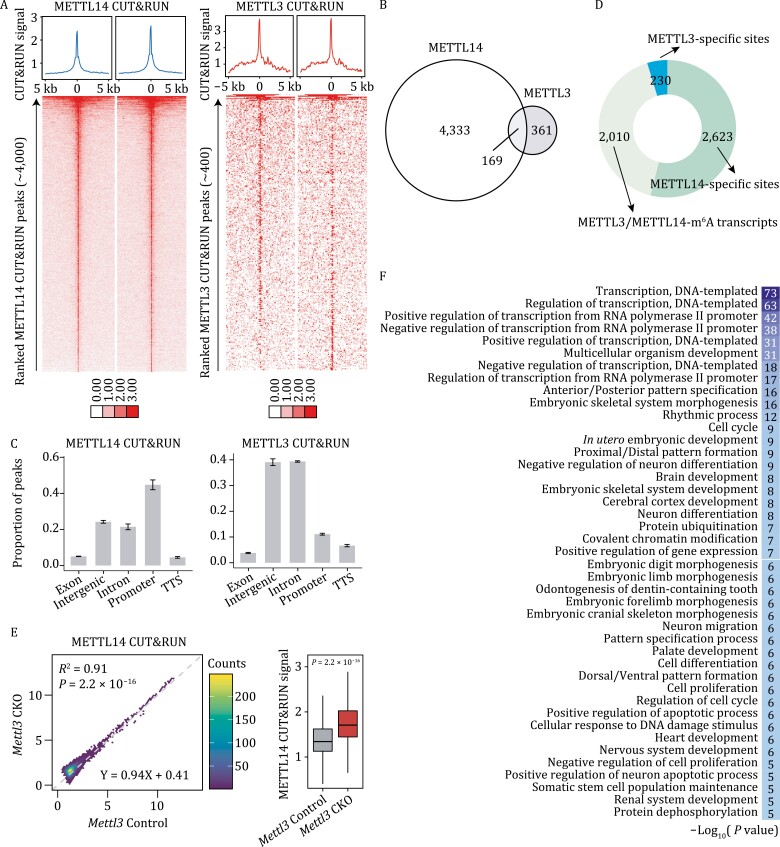
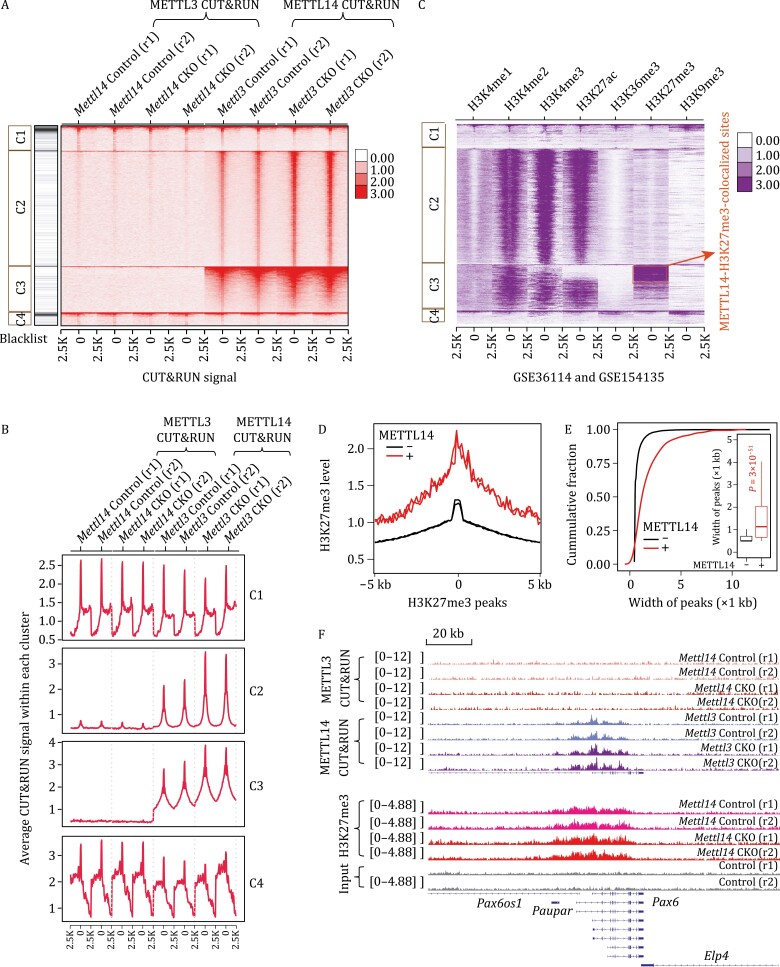
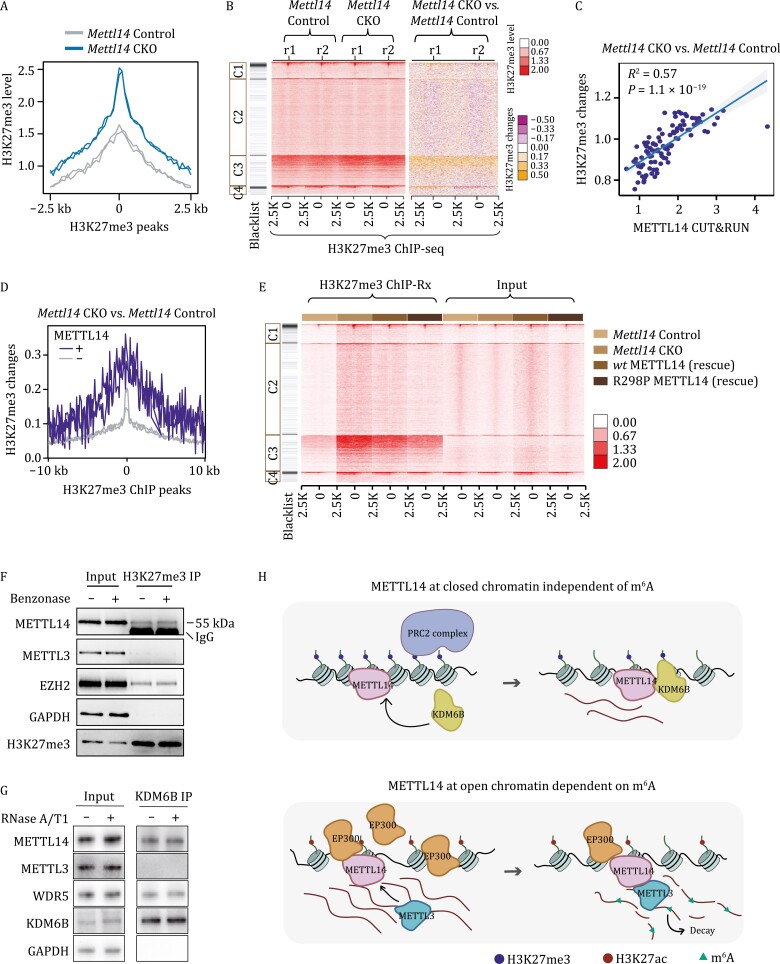
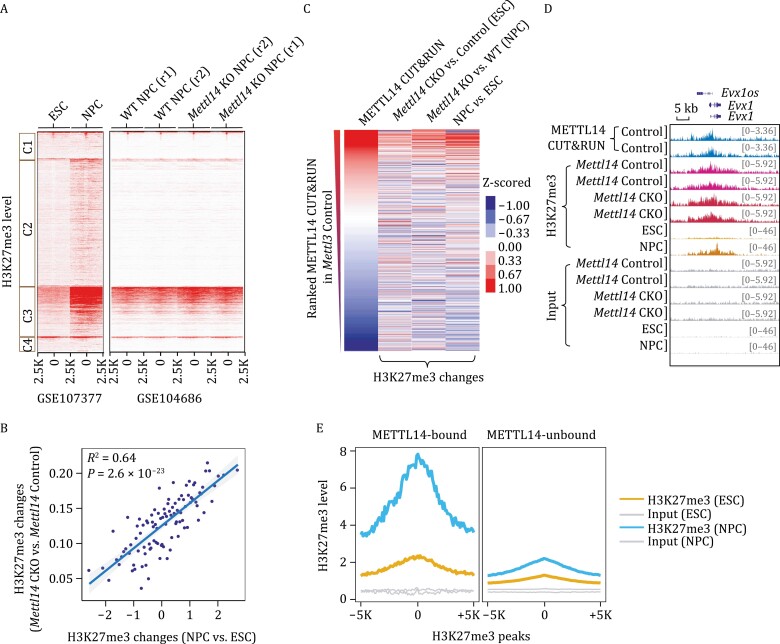
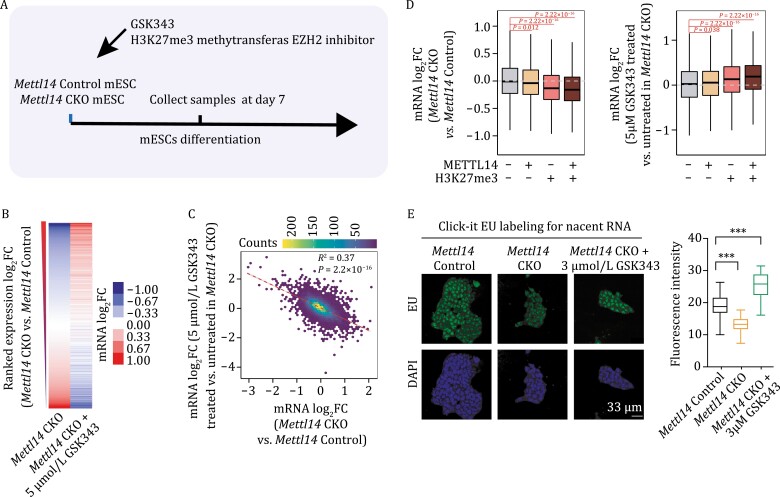
METTL3 and METTL14 are two components that form the core heterodimer of the main RNA m6A methyltransferase complex (MTC) that installs m6A. Surprisingly, depletion of METTL3 or METTL14 displayed distinct effects on stemness maintenance of mouse embryonic stem cell (mESC). While comparable global hypo-methylation in RNA m6A was observed in Mettl3 or Mettl14 knockout mESCs, respectively. Mettl14 knockout led to a globally decreased nascent RNA synthesis, whereas Mettl3 depletion resulted in transcription upregulation, suggesting that METTL14 might possess an m6A-independent role in gene regulation. We found that METTL14 colocalizes with the repressive H3K27me3 modification. Mechanistically, METTL14, but not METTL3, binds H3K27me3 and recruits KDM6B to induce H3K27me3 demethylation independent of METTL3. Depletion of METTL14 thus led to a global increase in H3K27me3 level along with a global gene suppression. The effects of METTL14 on regulation of H3K27me3 is essential for the transition from self-renewal to differentiation of mESCs. This work reveals a regulatory mechanism on heterochromatin by METTL14 in a manner distinct from METTL3 and independently of m6A, and critically impacts transcriptional regulation, stemness maintenance, and differentiation of mESCs.
Keywords: H3K27me3; METTL14; chromatin; m6A-independent; mESC differentiation.
©The Author(s) 2023. Published by Oxford University Press on behalf of Higher Education Press.
Conflict of interest statement
C.H. is a scientific founder and a member of the scientific advisory board of Accent Therapeutics, Inc. D
Figures
Similar articles
-
METTL14 regulates chromatin bivalent domains in mouse embryonic stem cells.Cell Rep. 2023 Jun 27;42(6):112650. doi: 10.1016/j.celrep.2023.112650. Epub 2023 Jun 13. Cell Rep. 2023. PMID: 37314930
-
Analysis of METTL3 and METTL14 in hepatocellular carcinoma.Aging (Albany NY). 2020 Nov 6;12(21):21638-21659. doi: 10.18632/aging.103959. Epub 2020 Nov 6. Aging (Albany NY). 2020. PMID: 33159022 Free PMC article.
-
METTL3 regulates m6A in endometrioid epithelial ovarian cancer independently of METTl14 and WTAP.Cell Biol Int. 2020 Dec;44(12):2524-2531. doi: 10.1002/cbin.11459. Epub 2020 Sep 11. Cell Biol Int. 2020. PMID: 32869897
-
Insights into roles of METTL14 in tumors.Cell Prolif. 2022 Jan;55(1):e13168. doi: 10.1111/cpr.13168. Epub 2021 Dec 13. Cell Prolif. 2022. PMID: 34904301 Free PMC article. Review.
-
The role, mechanism, and application of RNA methyltransferase METTL14 in gastrointestinal cancer.Mol Cancer. 2022 Aug 16;21(1):163. doi: 10.1186/s12943-022-01634-5. Mol Cancer. 2022. PMID: 35974338 Free PMC article. Review.
Cited by
-
METTL14/miR-29c-3p axis drives aerobic glycolysis to promote triple-negative breast cancer progression though TRIM9-mediated PKM2 ubiquitination.J Cell Mol Med. 2024 Feb;28(3):e18112. doi: 10.1111/jcmm.18112. Epub 2024 Jan 23. J Cell Mol Med. 2024. PMID: 38263865 Free PMC article.
-
The therapeutic potential of RNA m(6)A in lung cancer.Cell Commun Signal. 2024 Dec 31;22(1):617. doi: 10.1186/s12964-024-01980-5. Cell Commun Signal. 2024. PMID: 39736743 Free PMC article. Review.
-
The m6A reader YTHDC2 regulates UVB-induced DNA damage repair and histone modification.Photochem Photobiol. 2024 Jul-Aug;100(4):1031-1040. doi: 10.1111/php.13904. Epub 2024 Jan 8. Photochem Photobiol. 2024. PMID: 38190286 Free PMC article.
-
METTL Family in Healthy and Disease.Mol Biomed. 2024 Aug 19;5(1):33. doi: 10.1186/s43556-024-00194-y. Mol Biomed. 2024. PMID: 39155349 Free PMC article. Review.
-
M6A RNA modification: focusing on non-small cell lung cancer progression, therapeutic strategies and challenges.Front Oncol. 2025 Jul 16;15:1622359. doi: 10.3389/fonc.2025.1622359. eCollection 2025. Front Oncol. 2025. PMID: 40740874 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
