

CD8+ T cells promote HIV latency by remodeling CD4+ T cell metabolism to enhance their survival, quiescence, and stemness
- PMID: 37030290
- PMCID: PMC10880039
- DOI: 10.1016/j.immuni.2023.03.010
CD8+ T cells promote HIV latency by remodeling CD4+ T cell metabolism to enhance their survival, quiescence, and stemness
Abstract
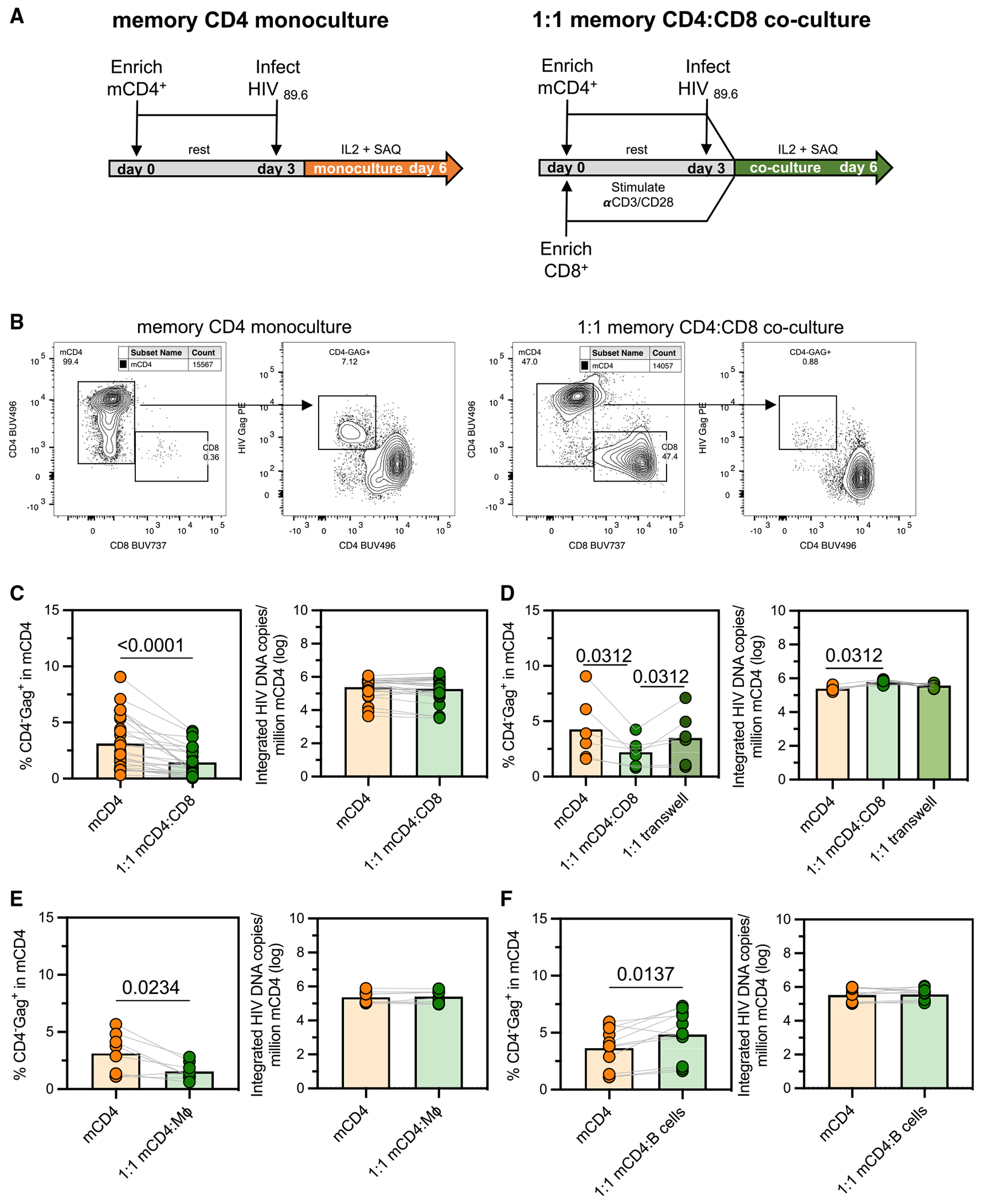
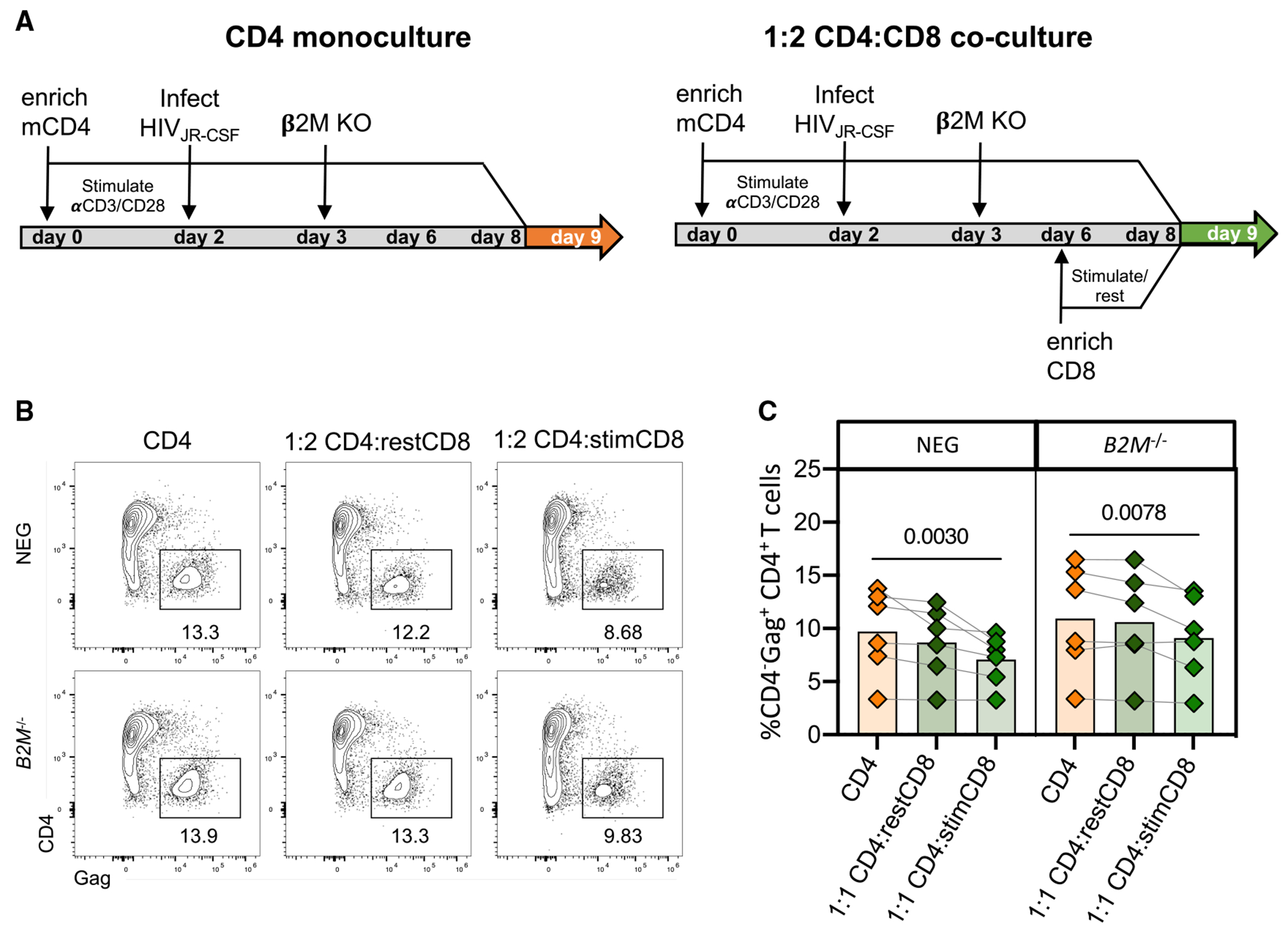
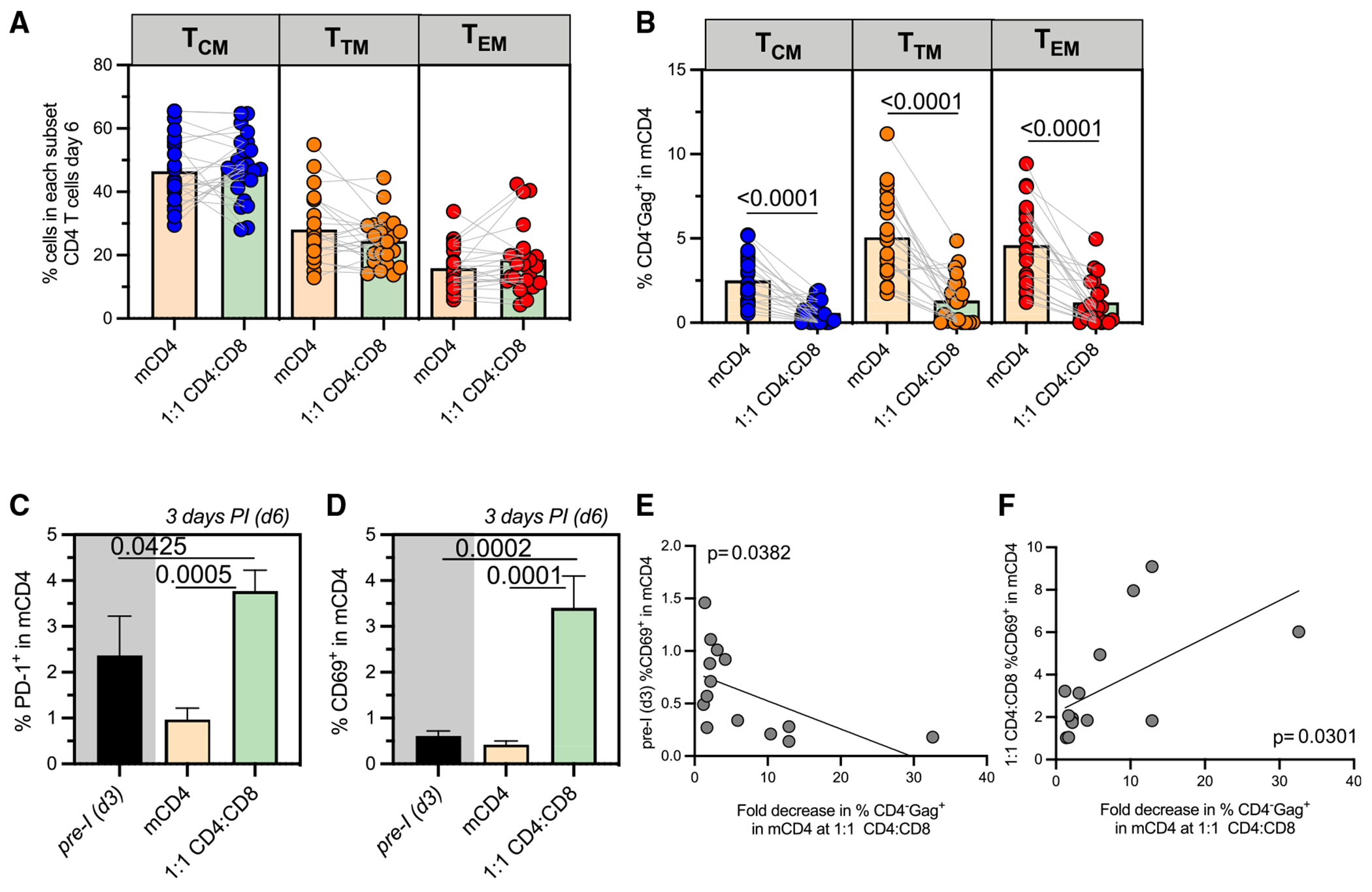
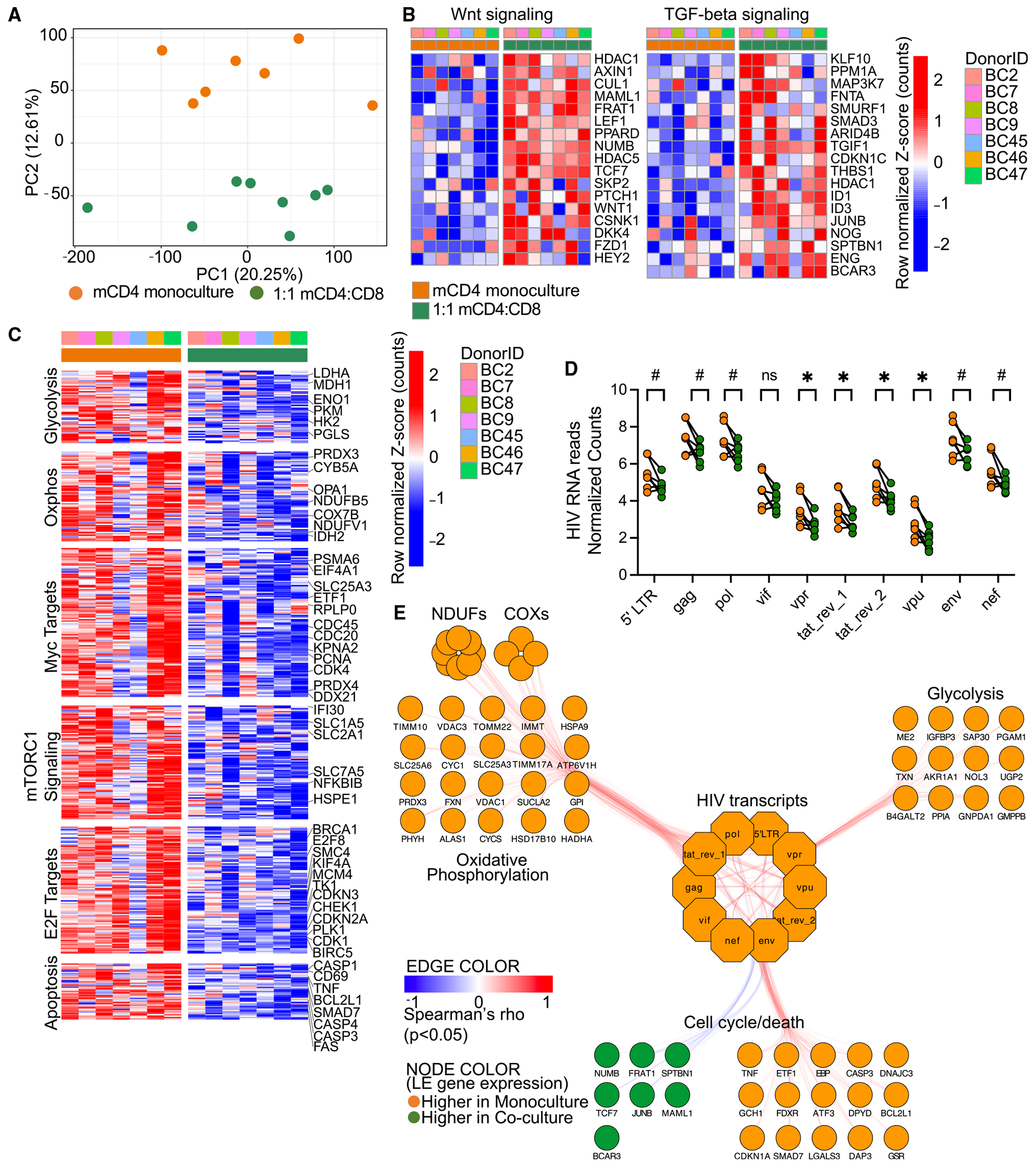
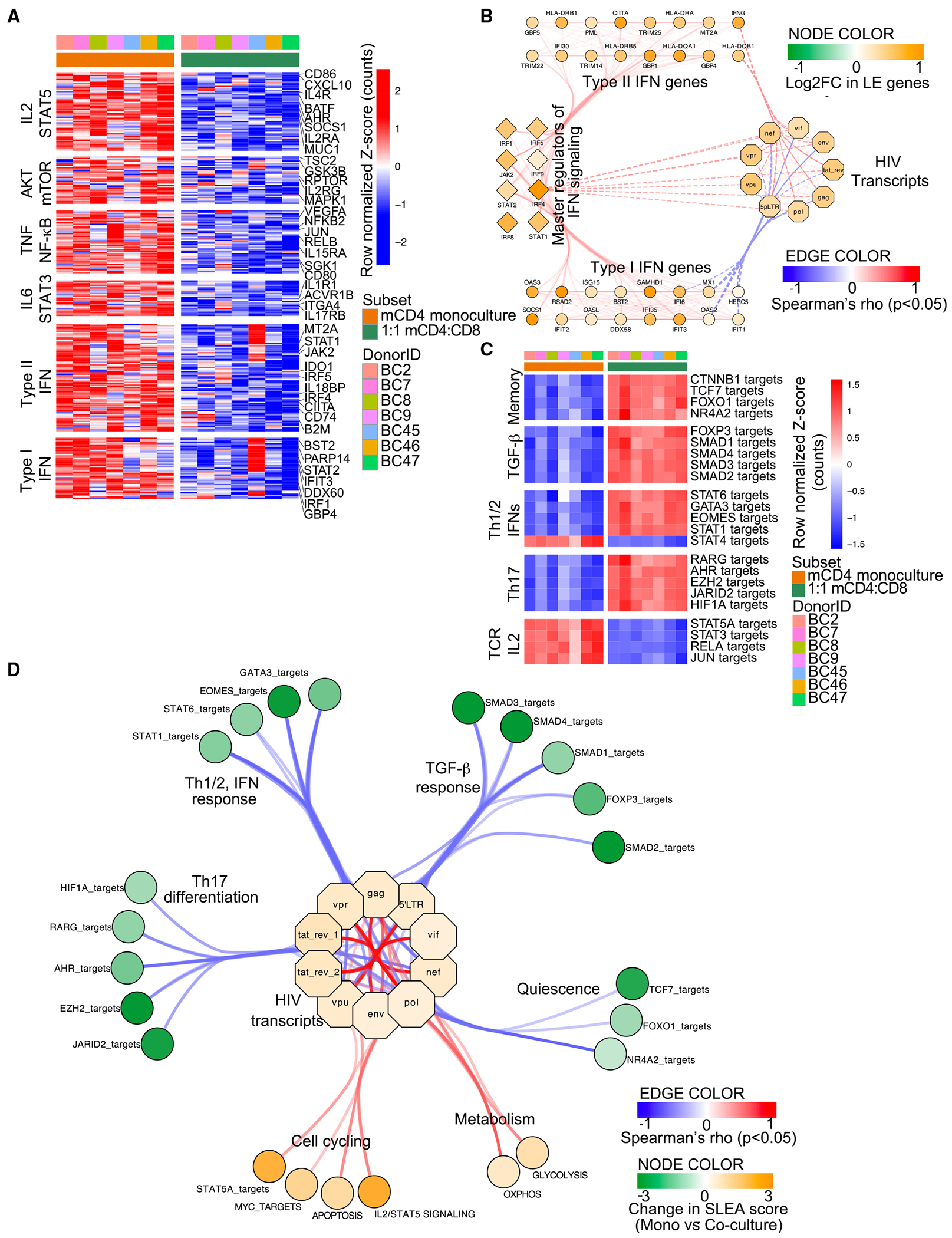
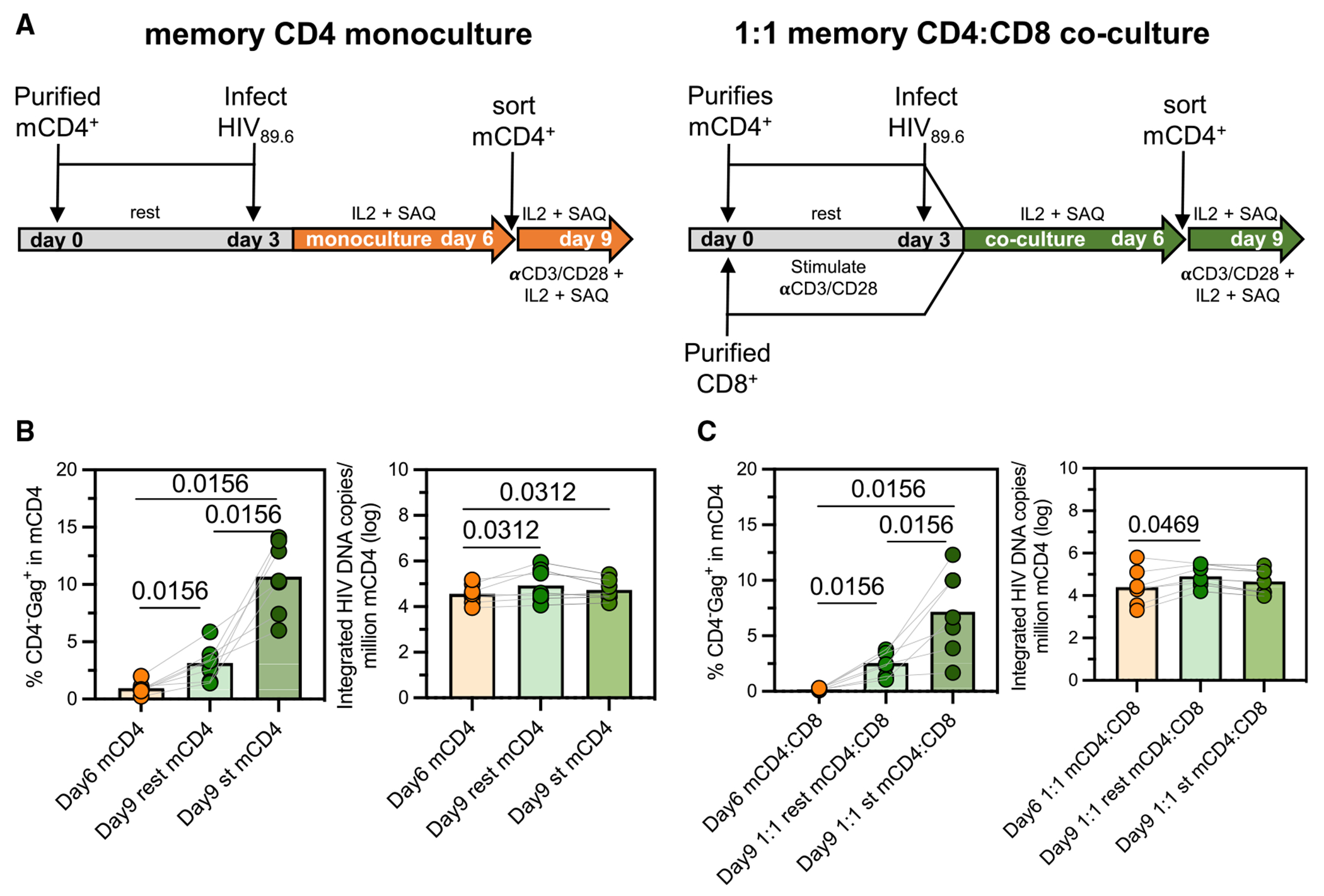
HIV infection persists during antiretroviral therapy (ART) due to a reservoir of latently infected cells that harbor replication-competent virus and evade immunity. Previous ex vivo studies suggested that CD8+ T cells from people with HIV may suppress HIV expression via non-cytolytic mechanisms, but the mechanisms responsible for this effect remain unclear. Here, we used a primary cell-based in vitro latency model and demonstrated that co-culture of autologous activated CD8+ T cells with HIV-infected memory CD4+ T cells promoted specific changes in metabolic and/or signaling pathways resulting in increased CD4+ T cell survival, quiescence, and stemness. Collectively, these pathways negatively regulated HIV expression and ultimately promoted the establishment of latency. As shown previously, we observed that macrophages, but not B cells, promoted latency in CD4+ T cells. The identification of CD8-specific mechanisms of pro-latency activity may favor the development of approaches to eliminate the viral reservoir in people with HIV.
Keywords: CD8 T cells; CD8 suppression; HIV; HIV cure; HIV latency; HIV reservoir; T cell biology.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Strategies for Management of Antiretroviral Therapy (SMART) Study Group, El-Sadr WM, Lundgren J, Neaton JD, Gordin F, Abrams D, Arduino RC, Babiker A, Burman W, Clumeck N, et al. (2006). CD4+ count-guided interruption of antiretroviral treatment. N. Engl. J. Med 355, 2283–2296. 10.1056/NEJMoa062360. - DOI - PubMed
-
- Finzi D, Hermankova M, Pierson T, Carruth LM, Buck C, Chaisson RE, Quinn TC, Chadwick K, Margolick J, Brookmeyer R, et al. (1997). Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 278, 1295–1300. - PubMed
-
- Wong JK, Hezareh M, Günthard HF, Havlir DV, Ignacio CC, Spina CA, and Richman DD (1997). Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 273, 1291–1295. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
