

The Long Noncoding RNA Cardiac Mesoderm Enhancer-Associated Noncoding RNA (Carmn) Is a Critical Regulator of Gastrointestinal Smooth Muscle Contractile Function and Motility
- PMID: 37030336
- PMCID: PMC10330198
- DOI: 10.1053/j.gastro.2023.03.229
The Long Noncoding RNA Cardiac Mesoderm Enhancer-Associated Noncoding RNA (Carmn) Is a Critical Regulator of Gastrointestinal Smooth Muscle Contractile Function and Motility
Abstract
Background & aims: Visceral smooth muscle cells (SMCs) are an integral component of the gastrointestinal (GI) tract that regulate GI motility. SMC contraction is regulated by posttranslational signaling and the state of differentiation. Impaired SMC contraction is associated with significant morbidity and mortality, but the mechanisms regulating SMC-specific contractile gene expression, including the role of long noncoding RNAs (lncRNAs), remain largely unexplored. Herein, we reveal a critical role of Carmn (cardiac mesoderm enhancer-associated noncoding RNA), an SMC-specific lncRNA, in regulating visceral SMC phenotype and contractility of the GI tract.
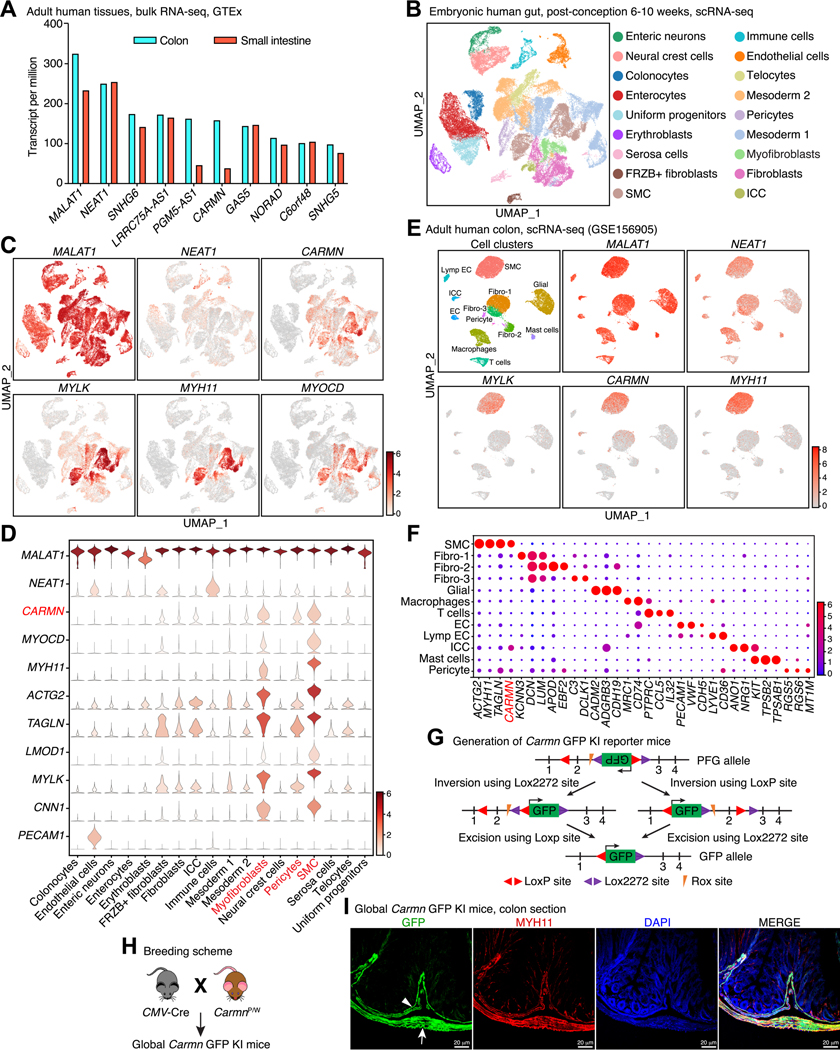
Methods: Genotype-Tissue Expression and publicly available single-cell RNA sequencing (scRNA-seq) data sets from embryonic, adult human, and mouse GI tissues were interrogated to identify SMC-specific lncRNAs. The functional role of Carmn was investigated using novel green fluorescent protein (GFP) knock-in (KI) reporter/knock-out (KO) mice. Bulk RNA-seq and single nucleus RNA sequencing (snRNA-seq) of colonic muscularis were used to investigate underlying mechanisms.
Results: Unbiased in silico analyses and GFP expression patterns in Carmn GFP KI mice revealed that Carmn is highly expressed in GI SMCs in humans and mice. Premature lethality was observed in global Carmn KO and inducible SMC-specific KO mice due to GI pseudo-obstruction and severe distension of the GI tract, with dysmotility in cecum and colon segments. Histology, GI transit, and muscle myography analysis revealed severe dilation, significantly delayed GI transit, and impaired GI contractility in Carmn KO vs control mice. Bulk RNA-seq of GI muscularis revealed that loss of Carmn promotes SMC phenotypic switching, as evidenced by up-regulation of extracellular matrix genes and down-regulation of SMC contractile genes, including Mylk, a key regulator of SMC contraction. snRNA-seq further revealed SMC Carmn KO not only compromised myogenic motility by reducing contractile gene expression but also impaired neurogenic motility by disrupting cell-cell connectivity in the colonic muscularis. These findings may have translational significance, because silencing CARMN in human colonic SMCs significantly attenuated contractile gene expression, including MYLK, and decreased SMC contractility. Luciferase reporter assays showed that CARMN enhances the transactivation activity of the master regulator of SMC contractile phenotype, myocardin, thereby maintaining the GI SMC myogenic program.
Conclusions: Our data suggest that Carmn is indispensable for maintaining GI SMC contractile function in mice and that loss of function of CARMN may contribute to human visceral myopathy. To our knowledge this is the first study showing an essential role of lncRNA in the regulation of visceral SMC phenotype.
Keywords: Carmn; Contractility; Pseudo-obstruction; Smooth Muscle; Visceral Myopathy; lncRNA.
Copyright © 2023 AGA Institute. All rights reserved.
Conflict of interest statement
Figures






Comment in
-
Sometimes Gut Smooth Muscle Forget That They Are Supposed to Contract: CARMN and Visceral Myopathy.Gastroenterology. 2023 Jul;165(1):27-29. doi: 10.1053/j.gastro.2023.05.003. Epub 2023 May 10. Gastroenterology. 2023. PMID: 37172742 Free PMC article. No abstract available.
References
-
- Sanders KM, Koh SD, Ward SM. Interstitial cells of cajal as pacemakers in the gastrointestinal tract. Annu Rev Physiol 2006;68:307–43. - PubMed
-
- Dong W, Baldwin C, Choi J, et al. Identification of a dominant MYH11 causal variant in chronic intestinal pseudo-obstruction: Results of whole-exome sequencing. Clin Genet 2019;96:473–477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
