

Development of mouse models with restricted HLA-B∗57:01 presentation for the study of flucloxacillin-driven T-cell activation and tolerance in liver injury
- PMID: 37030592
- PMCID: PMC10524621
- DOI: 10.1016/j.jaci.2023.03.029
Development of mouse models with restricted HLA-B∗57:01 presentation for the study of flucloxacillin-driven T-cell activation and tolerance in liver injury
Abstract
Background: Flucloxacillin (FLX)-induced liver injury is immune-mediated and highly associated to HLA-B∗57:01 expression. Host factors leading to drug-induced liver injury are not yet well understood.
Objective: Characterize in vivo immune mechanisms determining the development of CD8+ T cells reactive to FLX in animals expressing the risk human leukocyte antigen (HLA) allotype.
Methods: HLA-B∗57:01 transgenic mice (Tg) or Tg strains with H2-KbDb knockout (Tg/KO) or H2-KbDb/PD-1 double knockout (Tg/DKO) were treated with drug and/or anti-CD4 antibody. Drug-induced liver injury was evaluated on the basis of liver enzyme and histologic changes at day 10 of treatment. FLX-reactive CD8+ T cells were characterized in vitro by release of effector molecules on drug restimulation, gene expression, and flow cytometry analysis, and functionality tested for hepatic cytotoxicity.
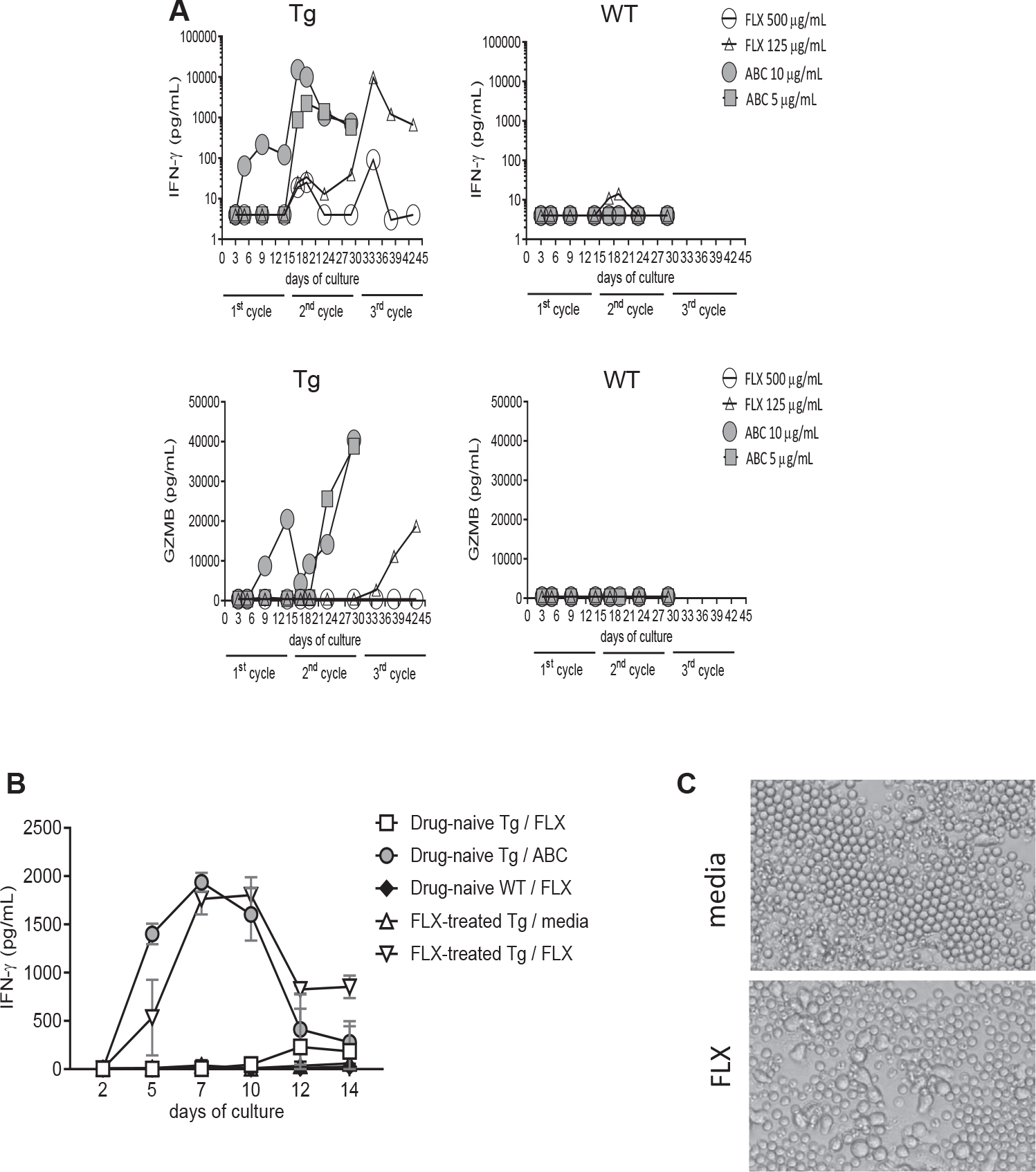
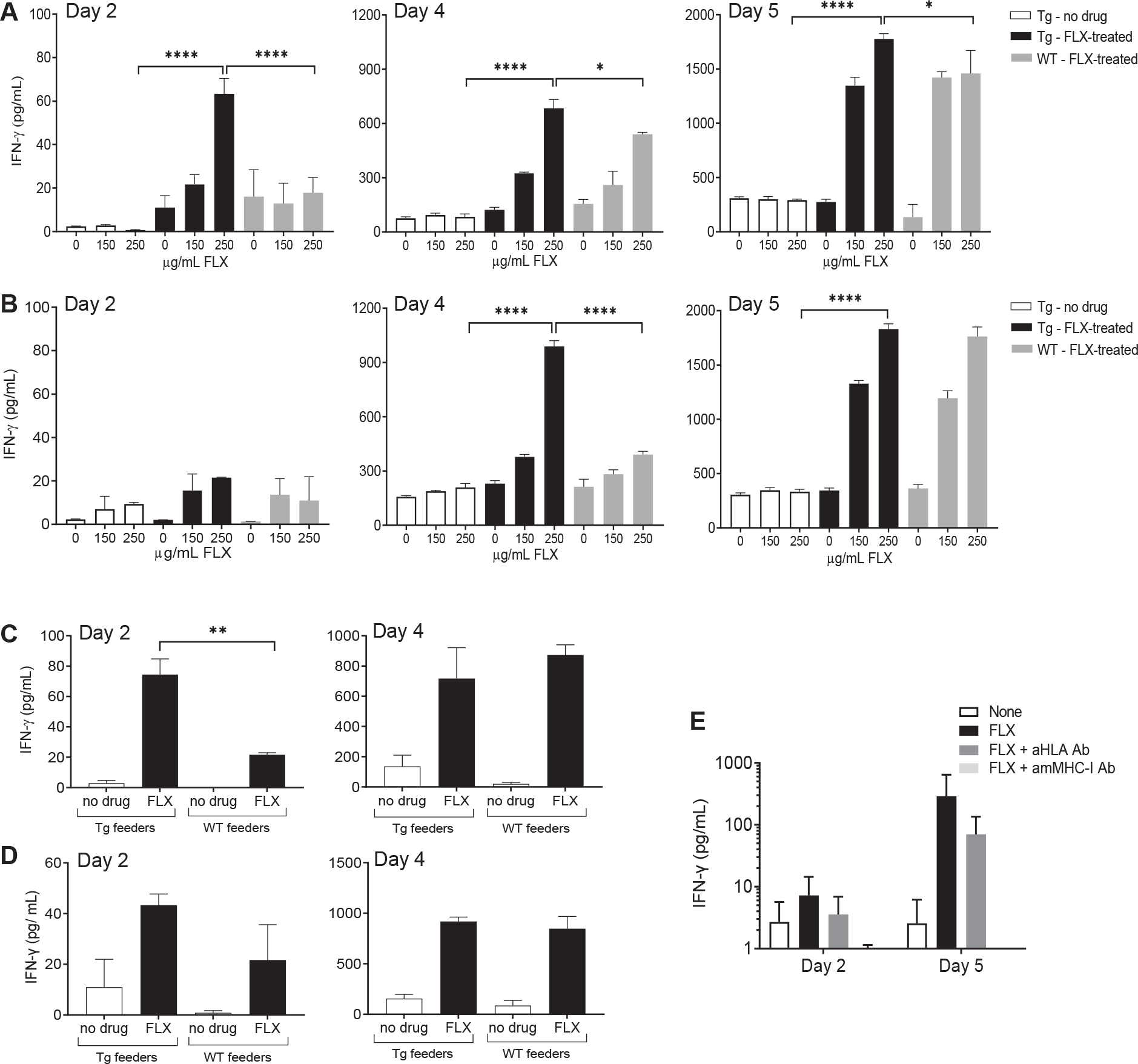
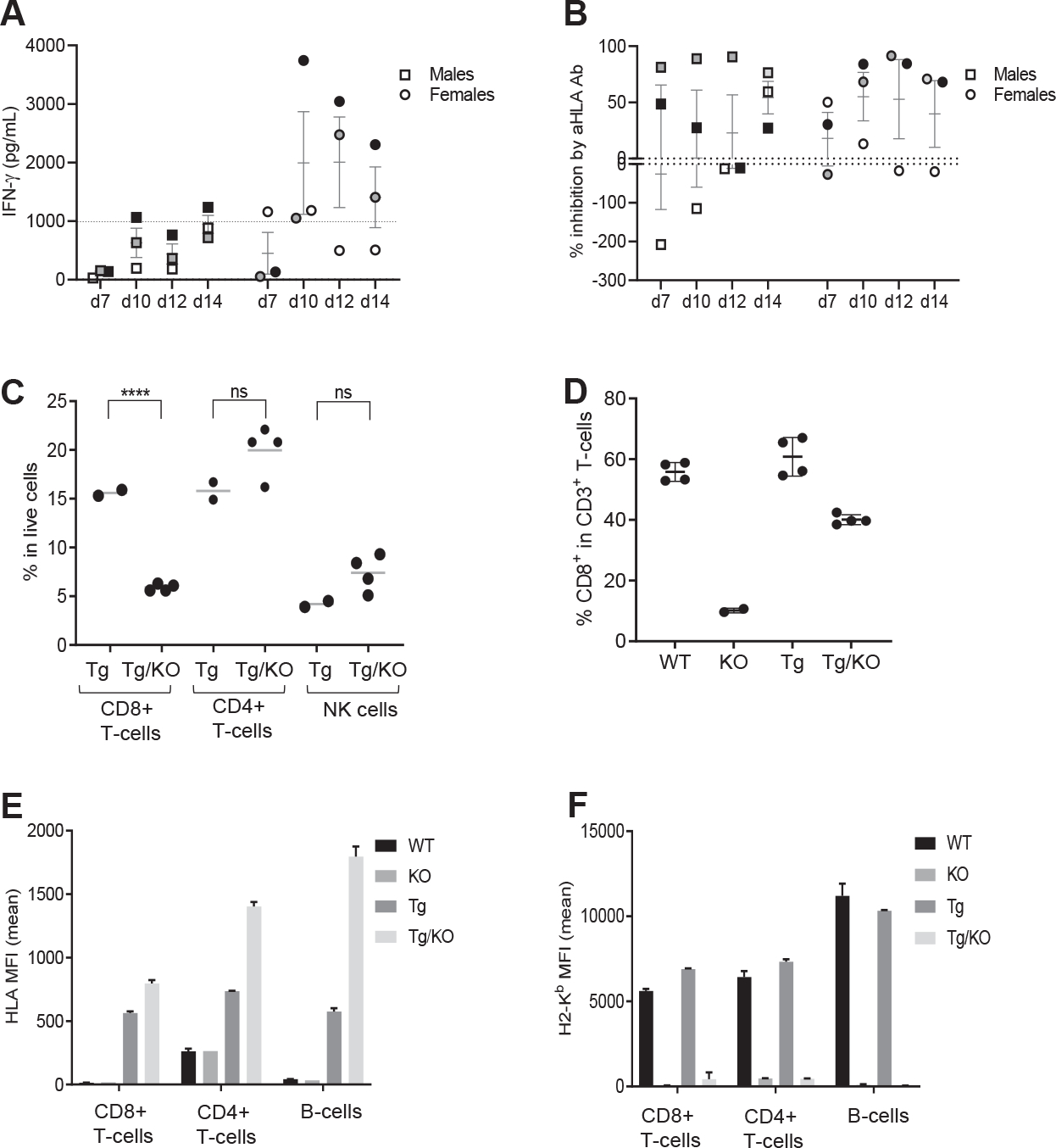
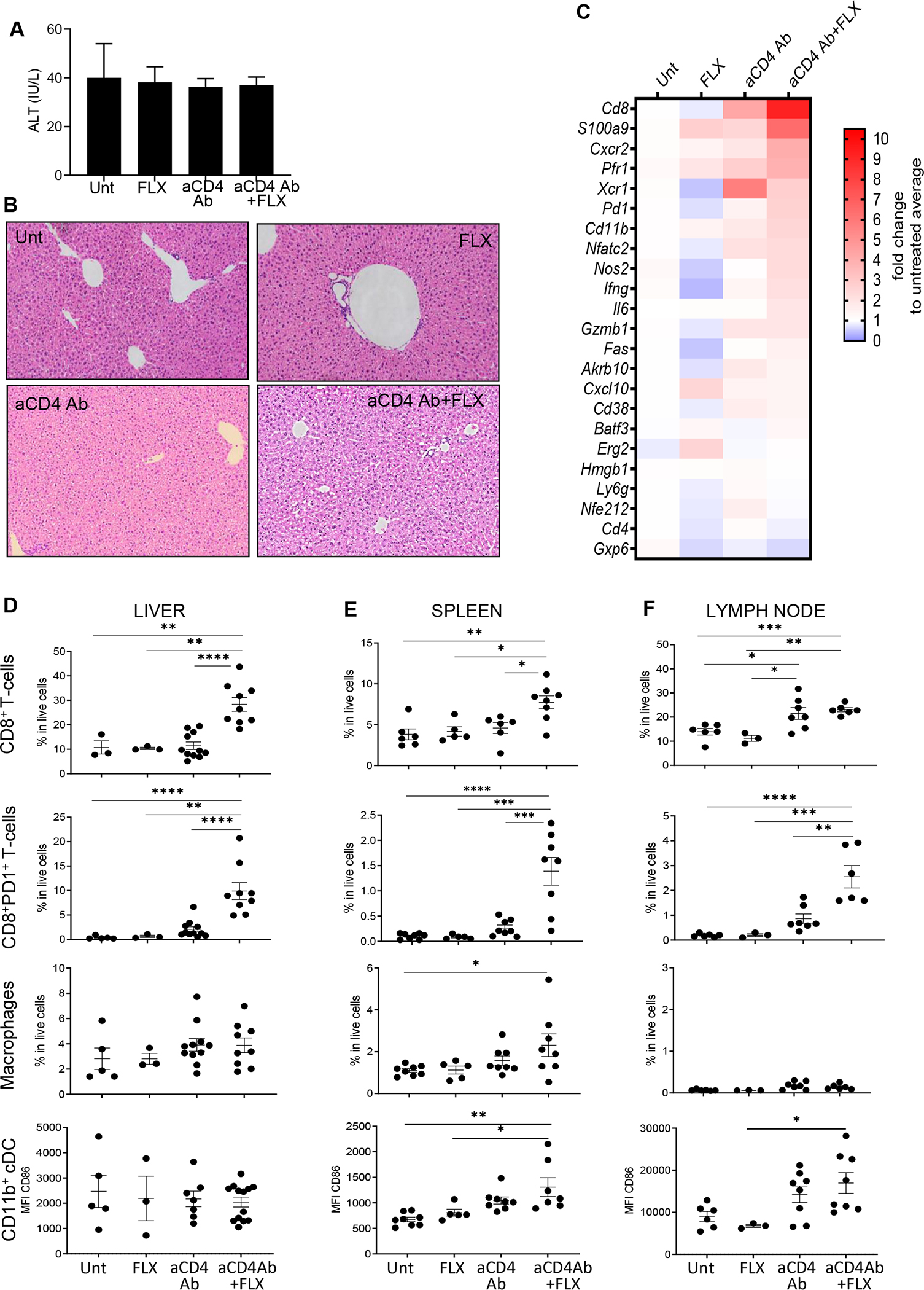
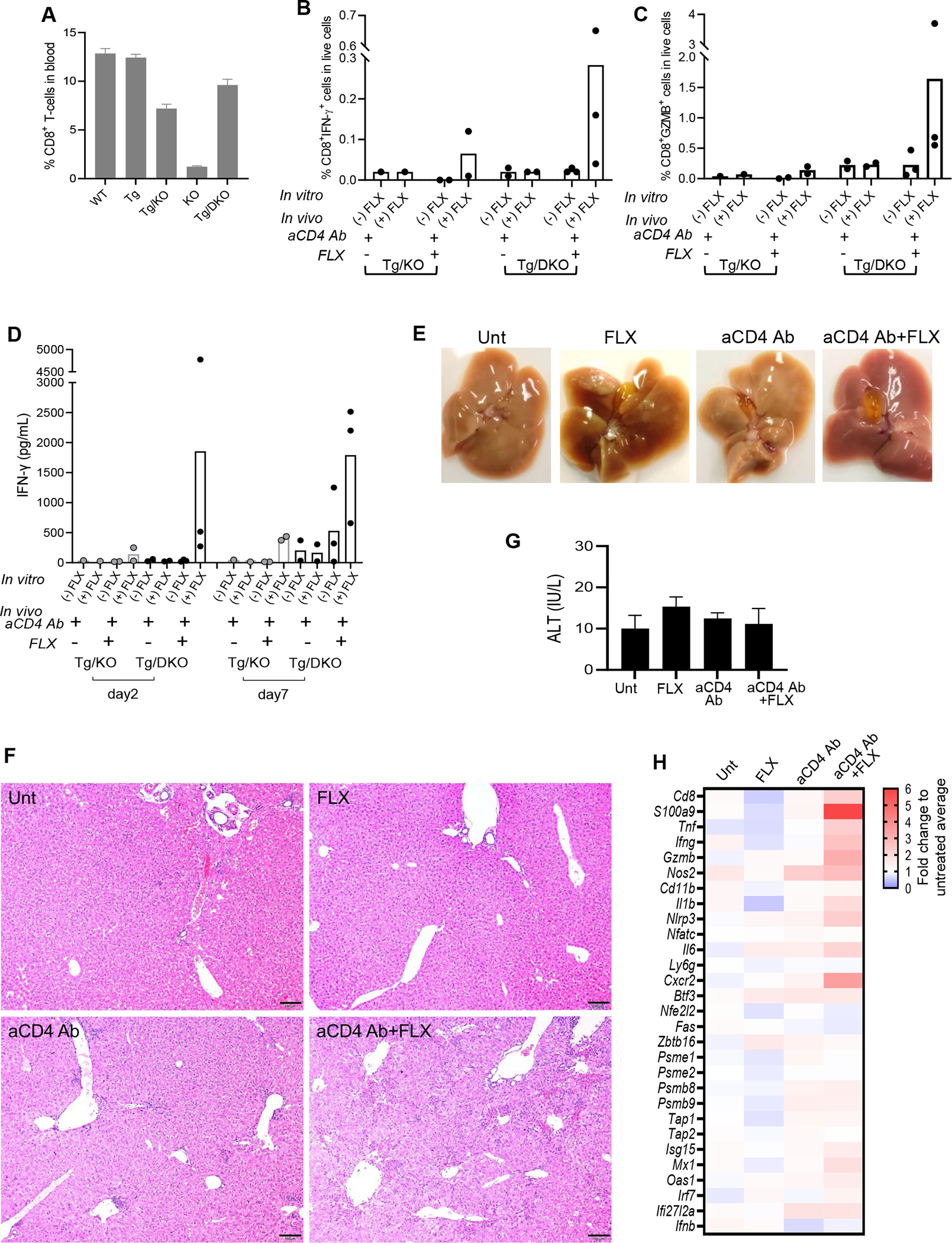
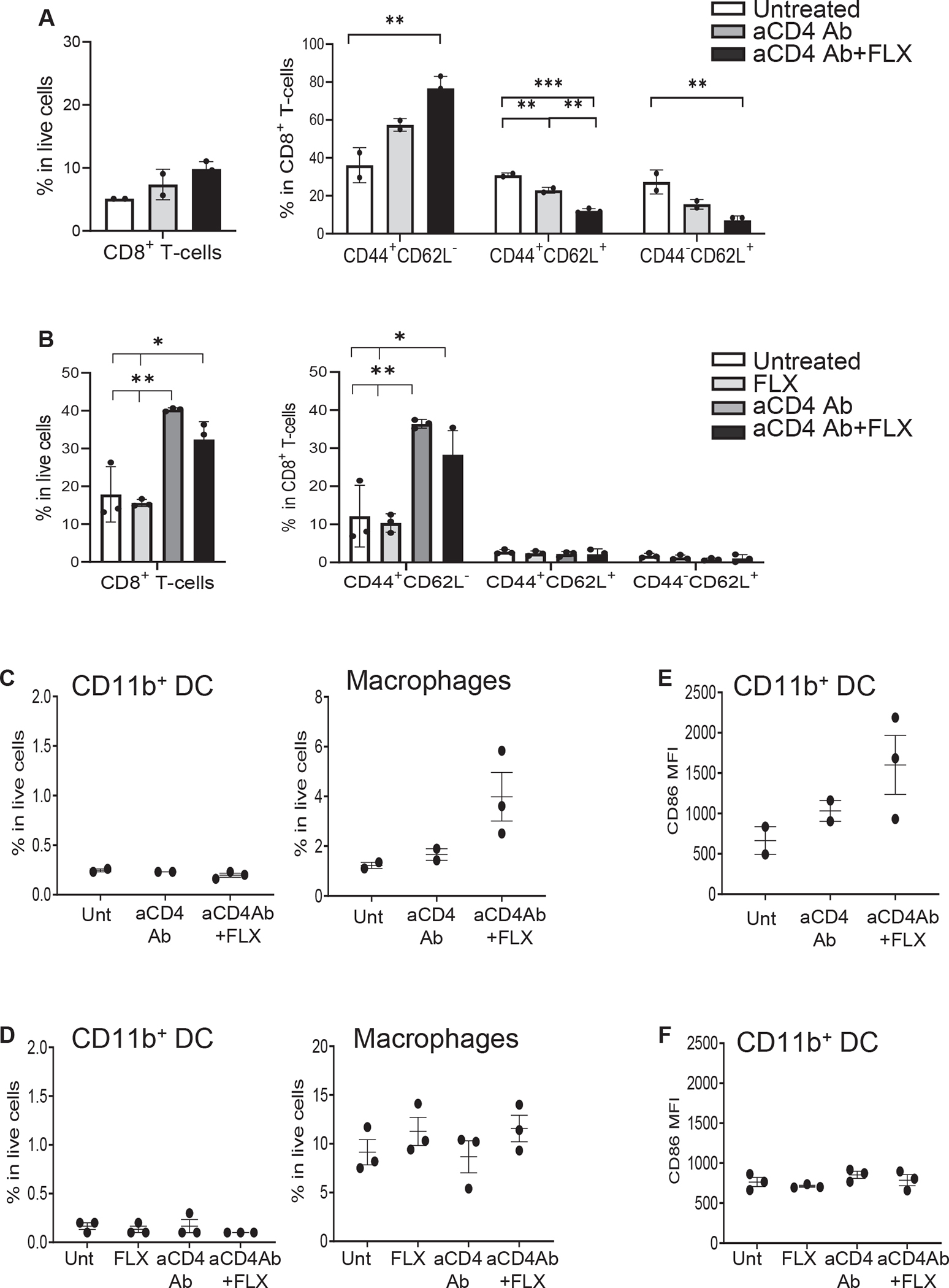
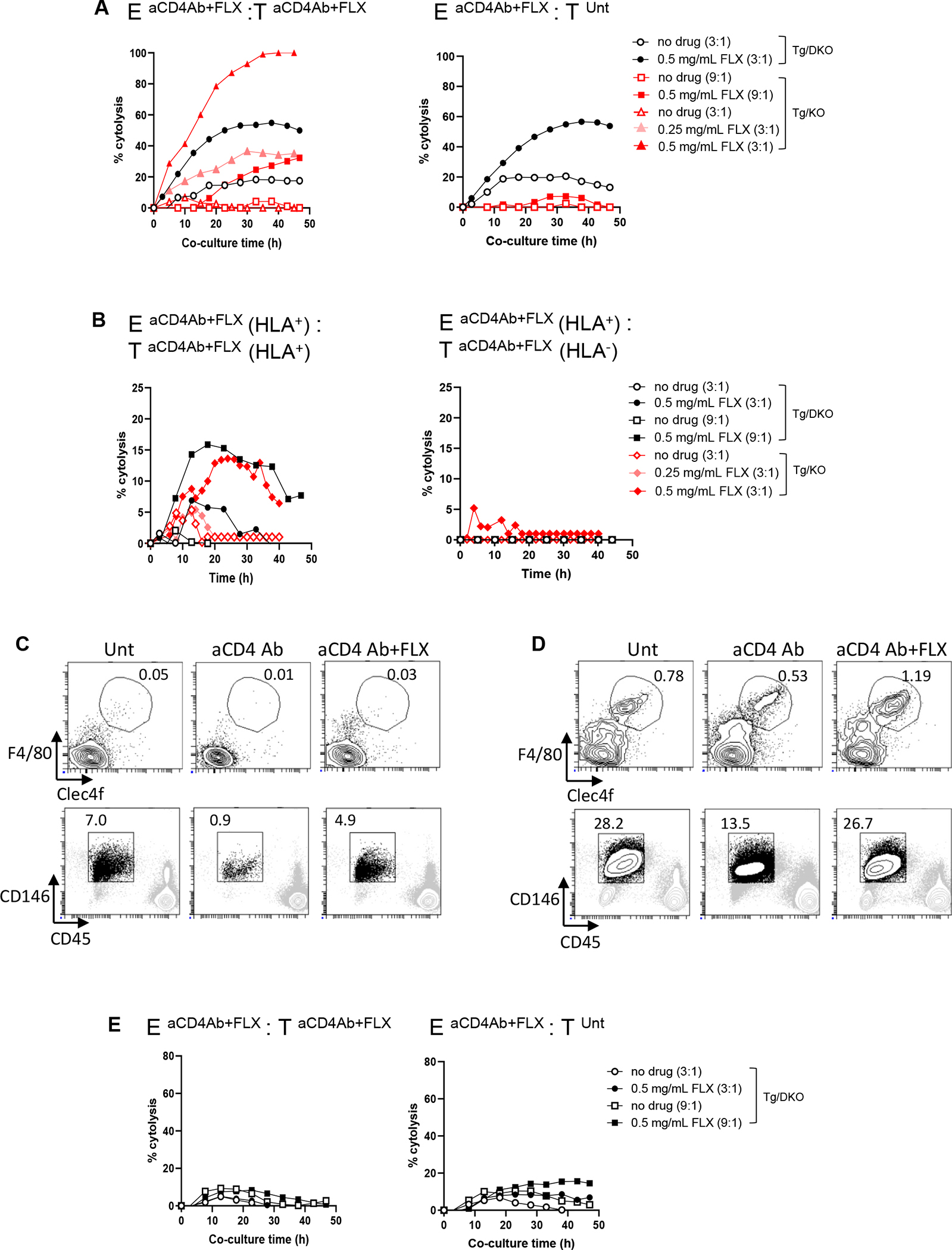
Results: CD8+ T-cell responses to FLX in Tg were dependent on both HLA and mouse major histocompatibility complex I presentation and in vivo priming. Eliminating H2-KbDb in Tg/KO to allow exclusive presentation of FLX by HLA resulted in a less robust drug-specific CD8+T-cell response unless CD4+ cells, including regulatory T cells, were depleted. Treatment of Tg/KO with anti-CD4 antibody and FLX led to subclinical liver inflammation associated with an increase in PD1+CD8+ T cells in the lymphoid organs and liver. Impaired PD-1 expression in Tg/DKO led to liver histopathologic and transcriptional alterations but without hepatic enzyme elevations. Moreover, effector lymphocytes accumulated in the liver and showed FLX-dependent hepatic cytotoxicity in vitro when tolerogenic liver cells were depleted.
Conclusions: In our in vivo models, FLX primes CD8+ T cells to recognize drug presented by HLA-B∗57:01 and murine major histocompatibility complex I. HLA-B∗57:01-dependent CD8+ T-cell reaction to FLX is limited by the presence of CD4+ cells, presumably regulatory T cells, and PD-1 expression. Tolerogenic hepatic cells limit clinical disease through PD-L1 or additional unexplored mechanisms.
Keywords: Flucloxacillin; HLA-B∗57:01 transgenic mice; drug-induced liver injury; tolerance.
Published by Elsevier Inc.
Conflict of interest statement
Disclosure of potential conflict of interest: The authors declare that they have no relevant conflicts of interest.
Figures
References
-
- Goh SJR, Tuomisto JEE, Purcell AW, Mifsud NA, Illing PT. The complexity of T cell–mediated penicillin hypersensitivity reactions. Allergy 2021;76:150–67. - PubMed
-
- Olsson R, Wiholm BE, Sand C, Zettergren L, Hultcrantz R, Myrhed M. Liver damage from flucloxacillin, cloxacillin and dicloxacillin. J Hepatol 1992;15:154–61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
