

Gamma secretase activity modulates BMP-7-induced dendritic growth in primary rat sympathetic neurons
- PMID: 37031474
- PMCID: PMC10330319
- DOI: 10.1016/j.autneu.2023.103085
Gamma secretase activity modulates BMP-7-induced dendritic growth in primary rat sympathetic neurons
Abstract
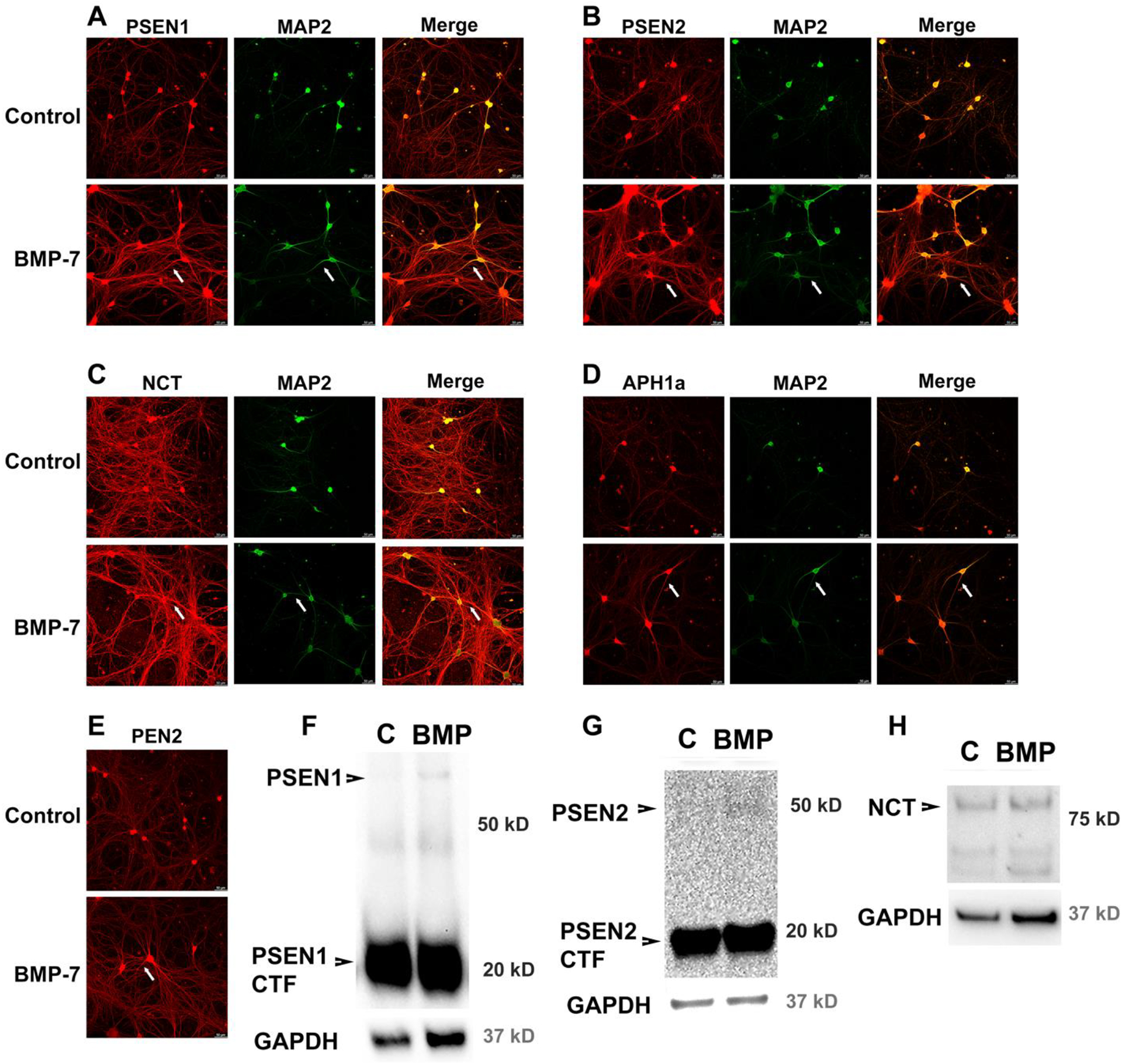
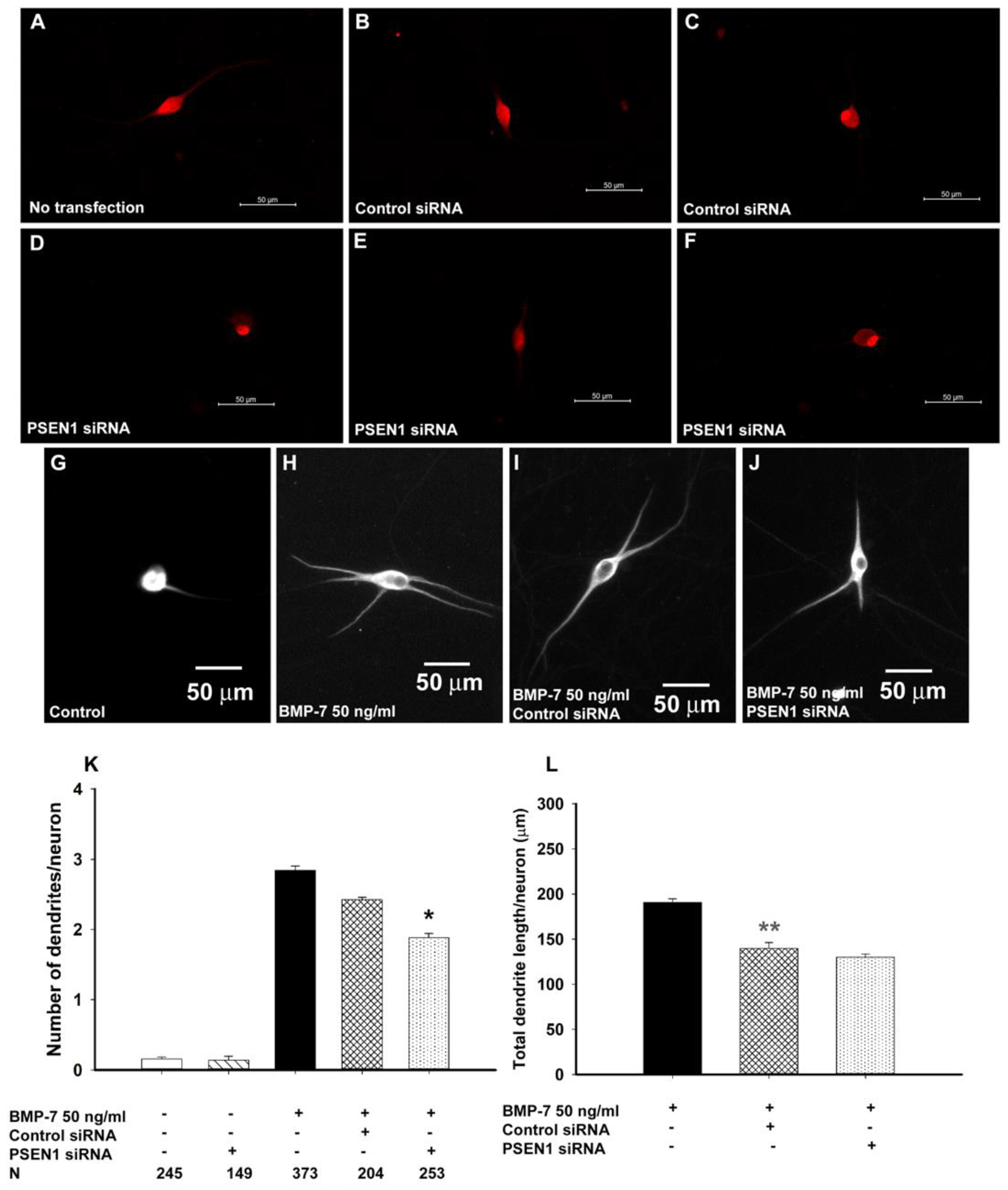
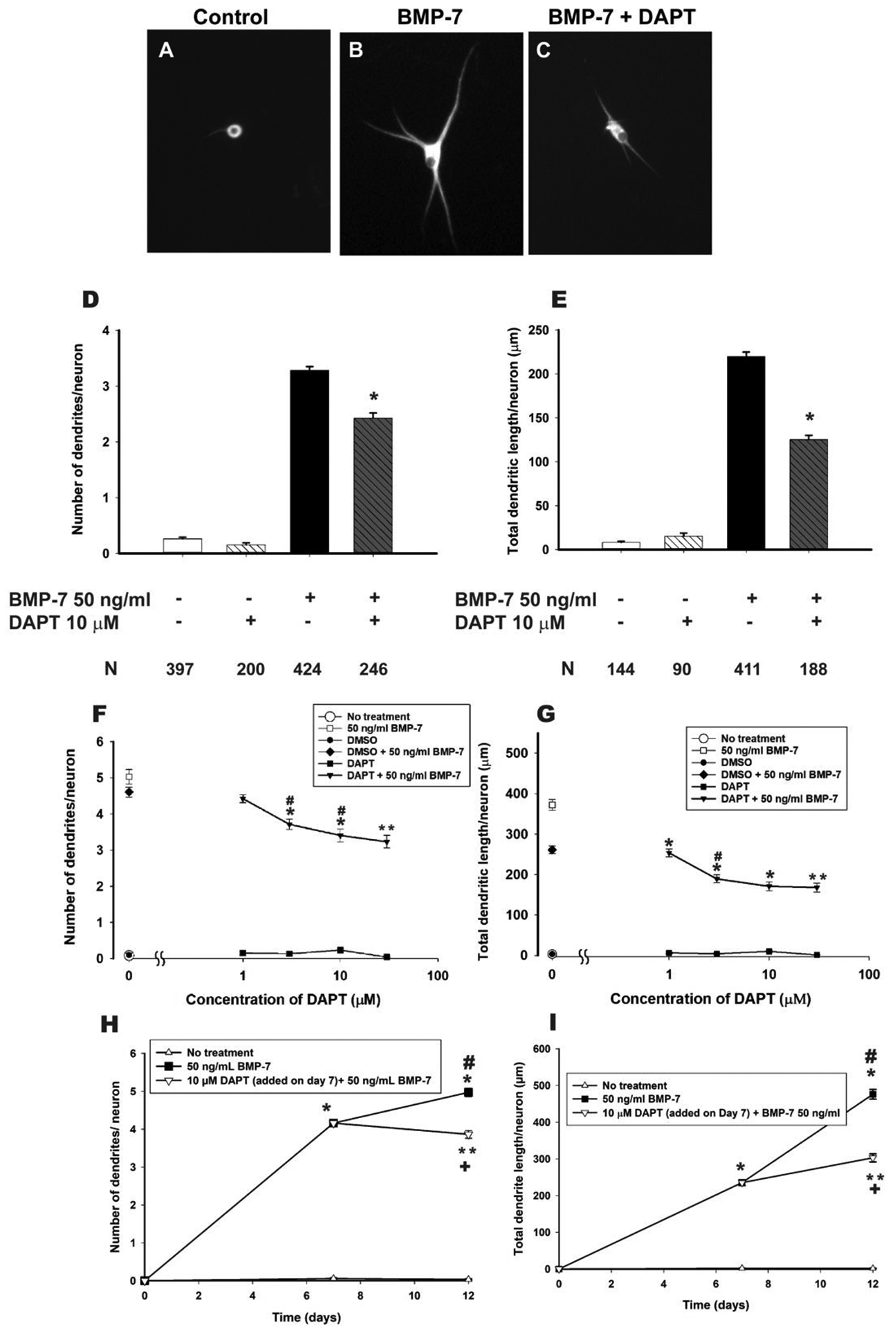
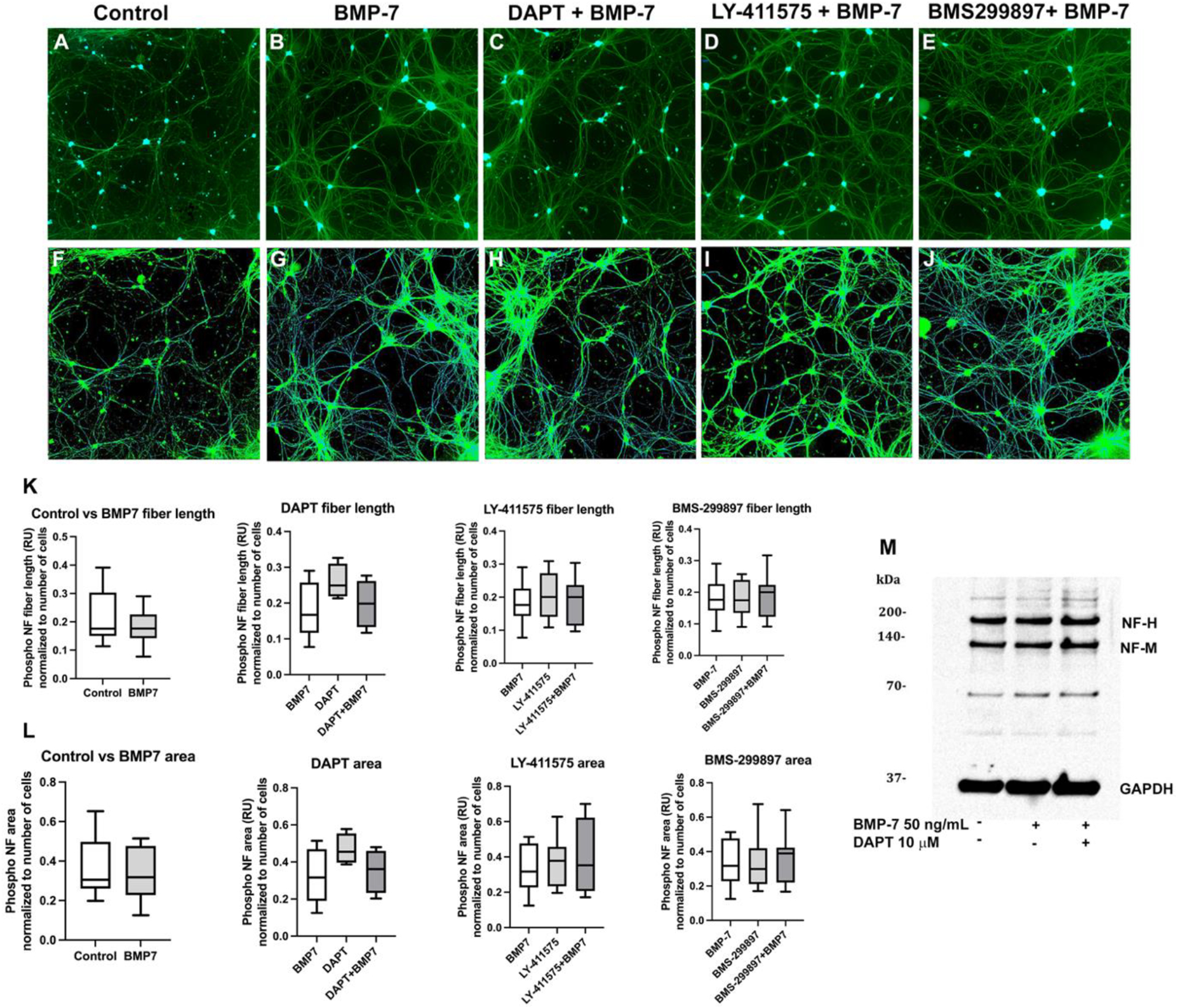
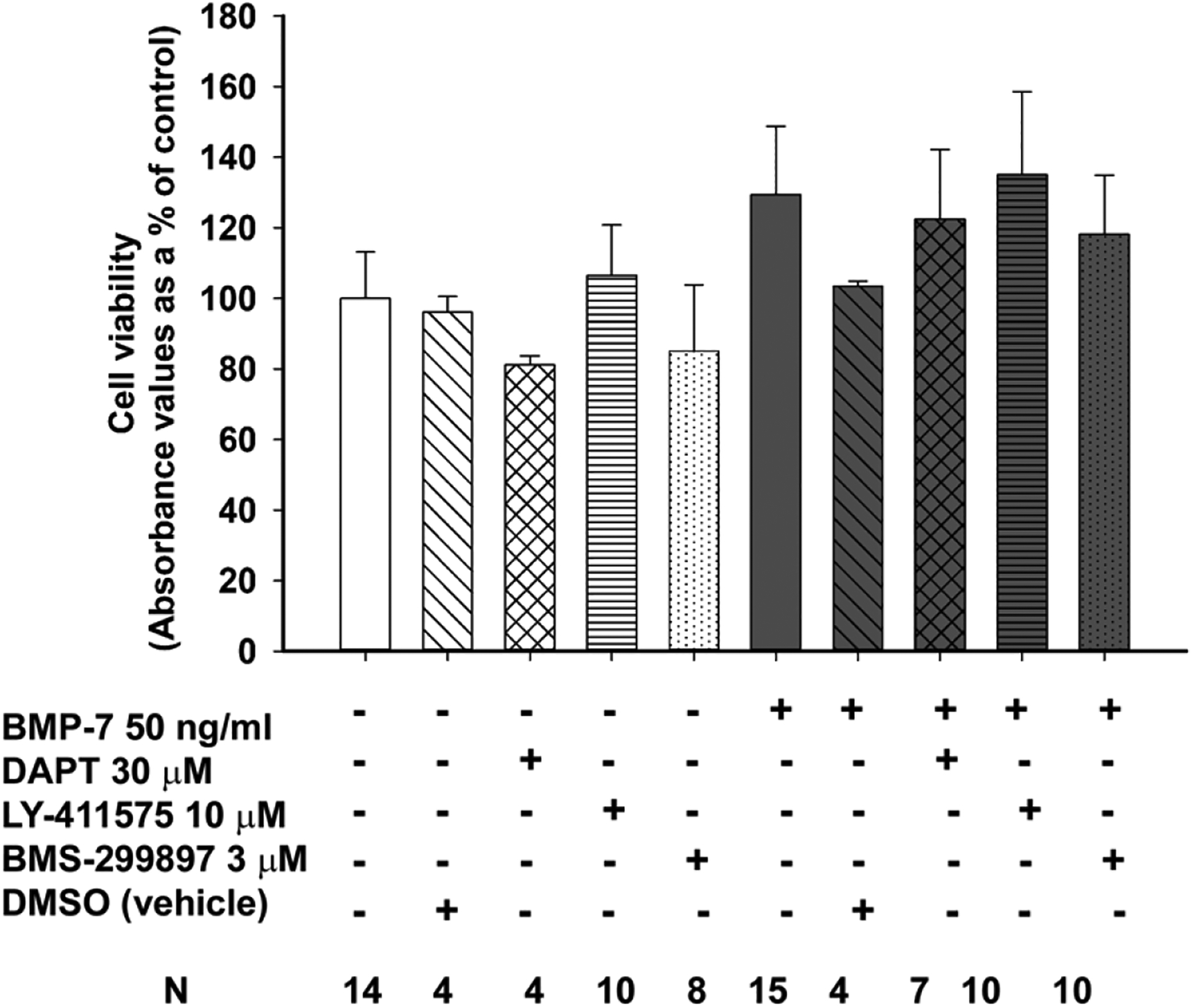
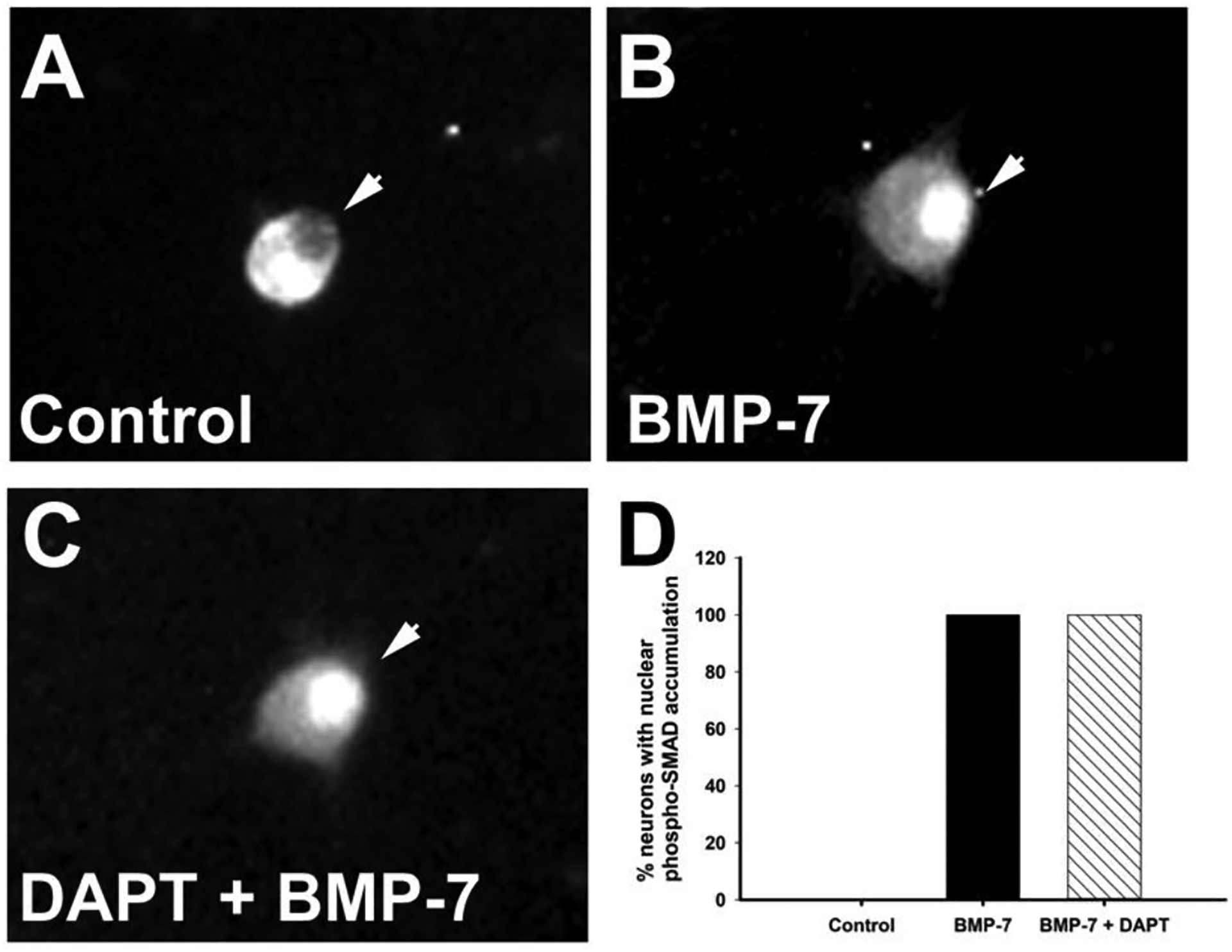
Autonomic dysfunction has been observed in Alzheimer's disease (AD); however, the effects of genes involved in AD on the peripheral nervous system are not well understood. Previous studies have shown that presenilin-1 (PSEN1), the catalytic subunit of the gamma secretase (γ-secretase) complex, mutations in which are associated with familial AD function, regulates dendritic growth in hippocampal neurons. In this study, we examined whether the γ-secretase pathway also influences dendritic growth in primary sympathetic neurons. Using immunoblotting and immunocytochemistry, molecules of the γ-secretase complex, PSEN1, PSEN2, PEN2, nicastrin and APH1a, were detected in sympathetic neurons dissociated from embryonic (E20/21) rat sympathetic ganglia. Addition of bone morphogenetic protein-7 (BMP-7), which induces dendrites in these neurons, did not alter expression or localization of γ-secretase complex proteins. BMP-7-induced dendritic growth was inhibited by siRNA knockdown of PSEN1 and by three γ-secretase inhibitors, γ-secretase inhibitor IX (DAPT), LY-411575 and BMS-299897. These effects were specific to dendrites and concentration-dependent and did not alter early downstream pathways of BMP signaling. In summary, our results indicate that γ-secretase activity enhances BMP-7 induced dendritic growth in sympathetic neurons. These findings provide insight into the normal cellular role of the γ-secretase complex in sympathetic neurons.
Keywords: BMS-299897; Bone morphogenetic proteins; DAPT; Dendrite; LY-411575; Presenilin; Sympathetic neurons; γ-Secretase.
Copyright © 2023 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors have no conflicts of interest to disclose.
Figures

Similar articles
-
MicroRNAs are Necessary for BMP-7-induced Dendritic Growth in Cultured Rat Sympathetic Neurons.Cell Mol Neurobiol. 2019 Oct;39(7):917-934. doi: 10.1007/s10571-019-00688-2. Epub 2019 May 18. Cell Mol Neurobiol. 2019. PMID: 31104181 Free PMC article.
-
Reactive oxygen species are involved in BMP-induced dendritic growth in cultured rat sympathetic neurons.Mol Cell Neurosci. 2015 Jul;67:116-25. doi: 10.1016/j.mcn.2015.06.007. Epub 2015 Jun 14. Mol Cell Neurosci. 2015. PMID: 26079955 Free PMC article.
-
Bone morphogenetic protein-5 (BMP-5) promotes dendritic growth in cultured sympathetic neurons.BMC Neurosci. 2001;2:12. doi: 10.1186/1471-2202-2-12. Epub 2001 Sep 11. BMC Neurosci. 2001. PMID: 11580864 Free PMC article.
-
Evidence For and Against a Pathogenic Role of Reduced γ-Secretase Activity in Familial Alzheimer's Disease.J Alzheimers Dis. 2016 Apr 4;52(3):781-99. doi: 10.3233/JAD-151186. J Alzheimers Dis. 2016. PMID: 27060961 Review.
-
Contribution of the Presenilins in the cell biology, structure and function of γ-secretase.Semin Cell Dev Biol. 2020 Sep;105:12-26. doi: 10.1016/j.semcdb.2020.02.005. Epub 2020 Mar 4. Semin Cell Dev Biol. 2020. PMID: 32146031 Review.
Cited by
-
Molecular and cellular neurocardiology in heart disease.J Physiol. 2025 Mar;603(7):1689-1728. doi: 10.1113/JP284739. Epub 2024 May 22. J Physiol. 2025. PMID: 38778747 Review.
References
-
- Anderson JJ, Holtz G, Baskin PP, Turner M, Rowe B, Wang B, Kounnas MZ, Lamb BT, Barten D, Felsenstein K, McDonald I, Srinivasan K, Munoz B, Wagner SL, 2005. Reductions in beta-amyloid concentrations in vivo by the gamma-secretase inhibitors BMS-289948 and BMS-299897. Biochem Pharmacol 69, 689–698. 10.1016/J.BCP.2004.11.015 - DOI - PubMed
-
- Barten DM, Guss VL, Corsa JA, Loo A, Hansel SB, Zheng M, Munoz B, Srinivasan K, Wang B, Robertson BJ, Polson CT, Wang J, Roberts SB, Hendrick JP, Anderson JJ, Loy JK, Denton R, Verdoorn TA, Smith DW, Felsenstein KM, 2004. Dynamics of -Amyloid Reductions in Brain, Cerebrospinal Fluid, and Plasma of -Amyloid Precursor Protein Transgenic Mice Treated with a -Secretase Inhibitor. Journal of Pharmacology and Experimental Therapeutics 312, 635–643. 10.1124/jpet.104.075408 - DOI - PubMed
-
- Bittner T, Fuhrmann M, Burgold S, Jung CKE, Volbracht C, Steiner H, Mitteregger G, Kretzschmar HA, Haass C, Herms J, 2009. Gamma-secretase inhibition reduces spine density in vivo via an amyloid precursor protein-dependent pathway. J Neurosci 29, 10405–10409. 10.1523/JNEUROSCI.2288-09.2009 - DOI - PMC - PubMed
