

Pharmacological modulation of TSPO in microglia/macrophages and neurons in a chronic neurodegenerative model of prion disease
- PMID: 37032328
- PMCID: PMC10084680
- DOI: 10.1186/s12974-023-02769-y
Pharmacological modulation of TSPO in microglia/macrophages and neurons in a chronic neurodegenerative model of prion disease
Abstract
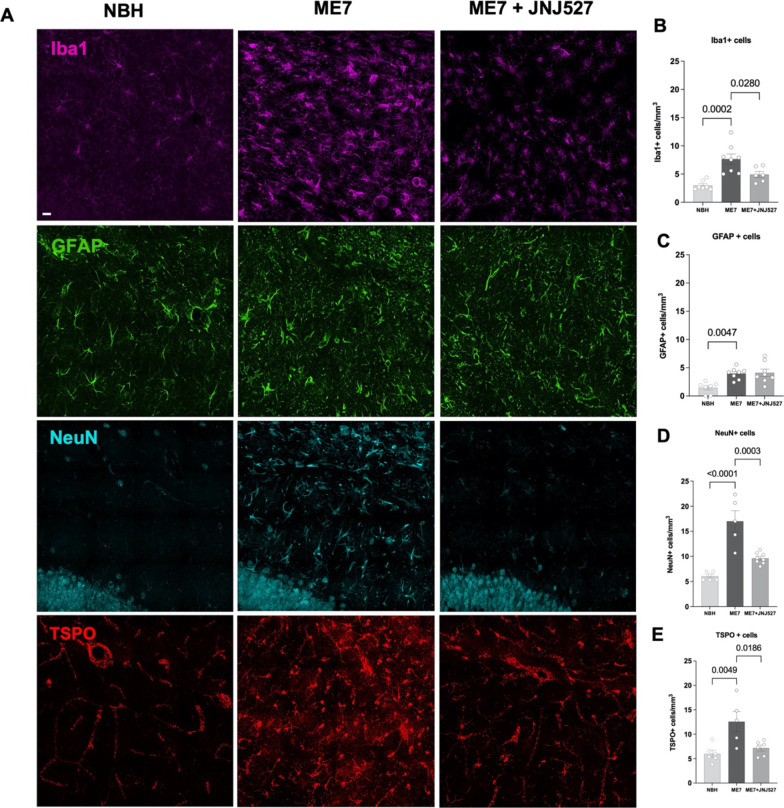
Neuroinflammation is an important component of many neurodegenerative diseases, whether as a primary cause or a secondary outcome. For that reason, either as diagnostic tools or to monitor progression and/or pharmacological interventions, there is a need for robust biomarkers of neuroinflammation in the brain. Mitochondrial TSPO (18 kDa Translocator protein) is one of few available biomarkers of neuroinflammation for which there are clinically available PET imaging agents. In this study, we further characterised neuroinflammation in a mouse model of prion-induced chronic neurodegeneration (ME7) including a pharmacological intervention via a CSF1R inhibitor. This was achieved by autoradiographic binding of the second-generation TSPO tracer, [3H]PBR28, along with a more comprehensive examination of the cellular contributors to the TSPO signal changes by immunohistochemistry. We observed regional increases of TSPO in the ME7 mouse brains, particularly in the hippocampus, cortex and thalamus. This increased TSPO signal was detected in the cells of microglia/macrophage lineage as well as in astrocytes, endothelial cells and neurons. Importantly, we show that the selective CSF1R inhibitor, JNJ-40346527 (JNJ527), attenuated the disease-dependent increase in TSPO signal, particularly in the dentate gyrus of the hippocampus, where JNJ527 attenuated the number of Iba1+ microglia and neurons, but not GFAP+ astrocytes or endothelial cells. These findings suggest that [3H]PBR28 quantitative autoradiography in combination with immunohistochemistry are important translational tools for detecting and quantifying neuroinflammation, and its treatments, in neurodegenerative disease. Furthermore, we demonstrate that although TSPO overexpression in the ME7 brains was driven by various cell types, the therapeutic effect of the CSF1R inhibitor was primarily to modulate TSPO expression in microglia and neurons, which identifies an important route of biological action of this particular CSF1R inhibitor and provides an example of a cell-specific effect of this type of therapeutic agent on the neuroinflammatory process.
Keywords: Astrocytes; CSF1R; ME7; Microglia; Neuroinflammation; Neurons; Prion disease; TSPO.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that there are no conflicts of financial and non-financial interests with respect to the conducted research, authorship and publication of this work.
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
