

Tunable control of insect pheromone biosynthesis in Nicotiana benthamiana
- PMID: 37032497
- PMCID: PMC10281601
- DOI: 10.1111/pbi.14048
Tunable control of insect pheromone biosynthesis in Nicotiana benthamiana
Abstract
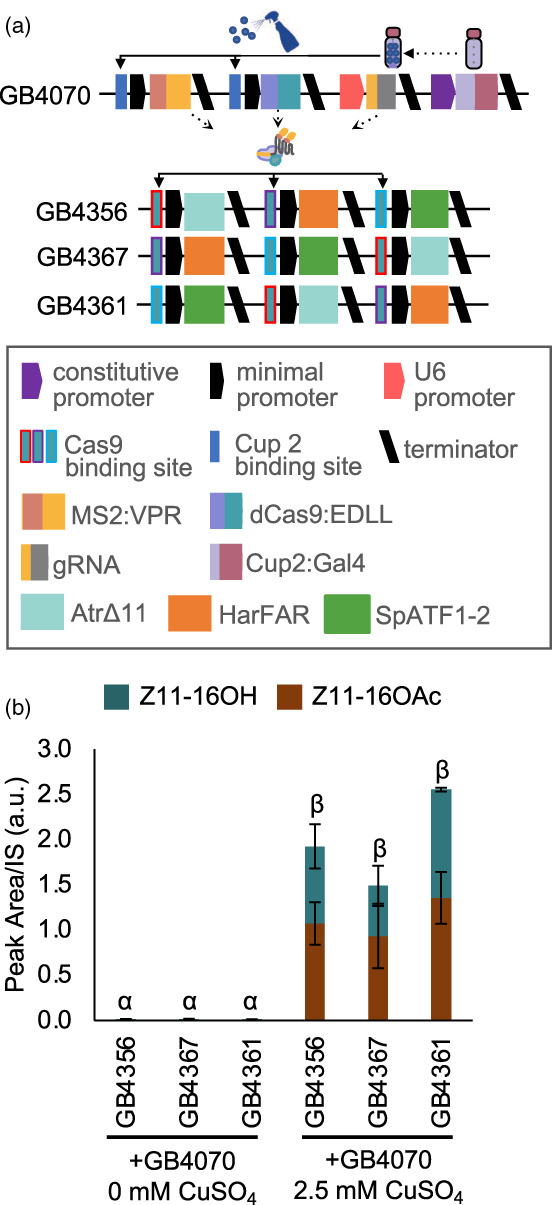
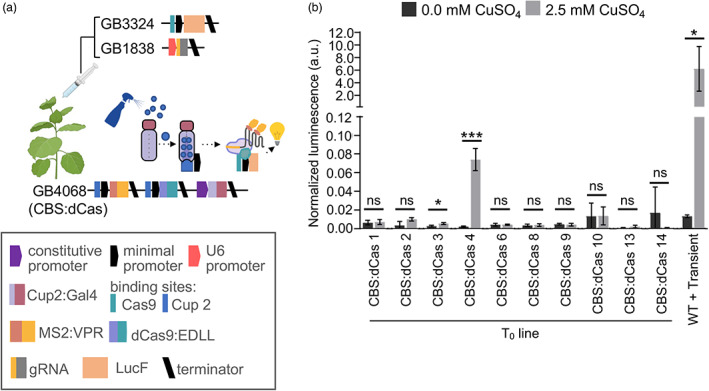
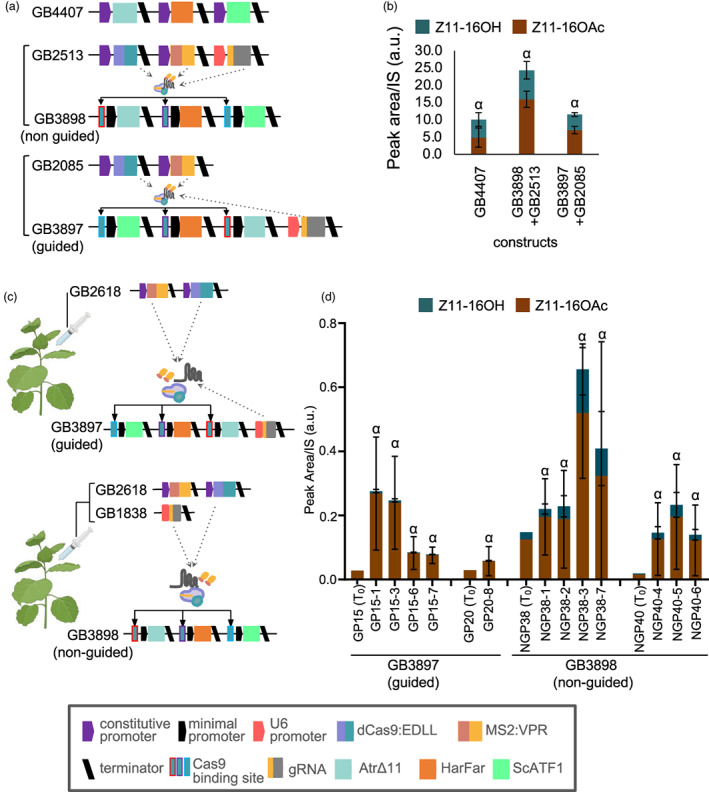
Previous work has demonstrated that plants can be used as production platforms for molecules used in health, medicine, and agriculture. Production has been exemplified in both stable transgenic plants and using transient expression strategies. In particular, species of Nicotiana have been engineered to produce a range of useful molecules, including insect sex pheromones, which are valued for species-specific control of agricultural pests. To date, most studies have relied on strong constitutive expression of all pathway genes. However, work in microbes has demonstrated that yields can be improved by controlling and balancing gene expression. Synthetic regulatory elements that provide control over the timing and levels of gene expression are therefore useful for maximizing yields from heterologous biosynthetic pathways. In this study, we demonstrate the use of pathway engineering and synthetic genetic elements for controlling the timing and levels of production of Lepidopteran sex pheromones in Nicotiana benthamiana. We demonstrate that copper can be used as a low-cost molecule for tightly regulated inducible expression. Further, we show how construct architecture influences relative gene expression and, consequently, product yields in multigene constructs. We compare a number of synthetic orthogonal regulatory elements and demonstrate maximal yields from constructs in which expression is mediated by dCas9-based synthetic transcriptional activators. The approaches demonstrated here provide new insights into the heterologous reconstruction of metabolic pathways in plants.
Keywords: Nicotiana benthamiana; metabolic engineering; pheromones; synthetic promoters; transcriptional activation.
© 2023 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
None declared.
Figures
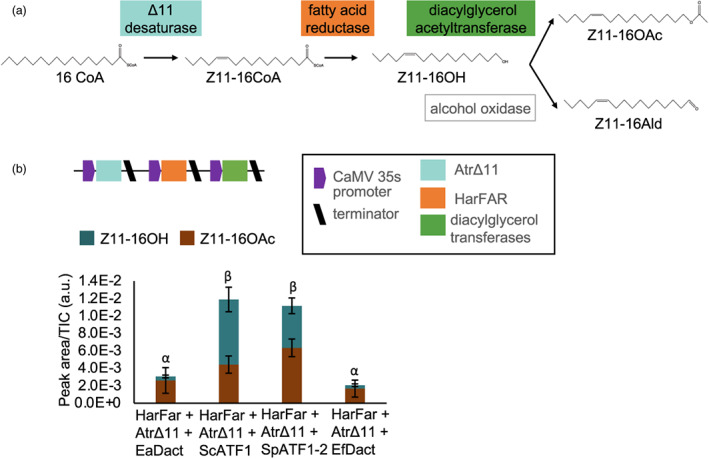
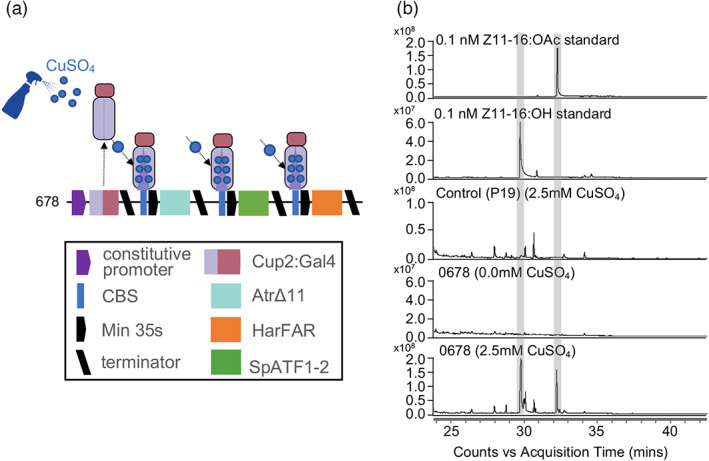
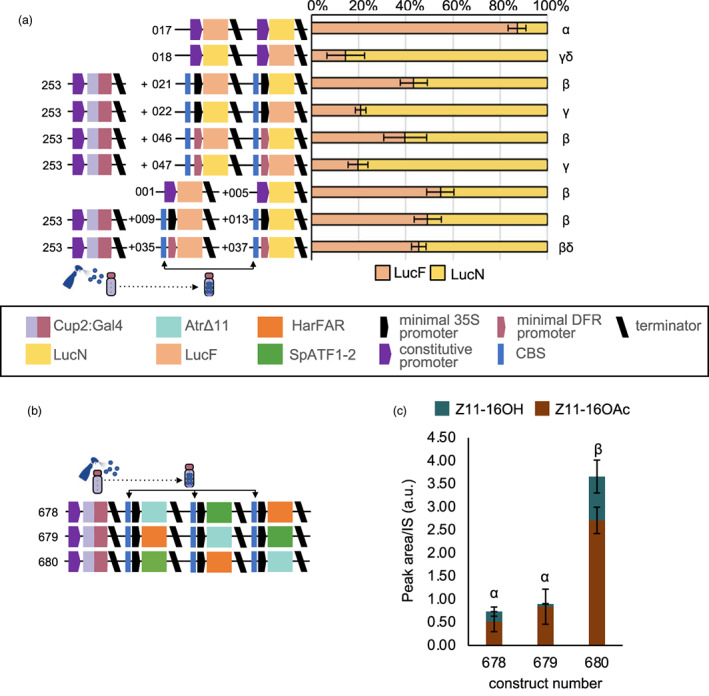
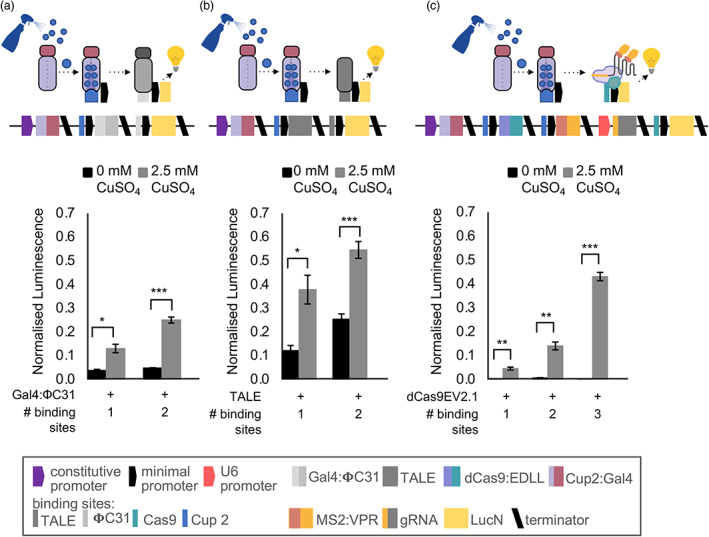
References
-
- Bally, J. , Jung, H. , Mortimer, C. , Naim, F. , Philips, J.G. , Hellens, R. , Bombarely, A. et al. (2018) The rise and rise of Nicotiana benthamiana: a plant for all reasons. Annu. Rev. Phytopathol. 56, 405–426. - PubMed
-
- Brodie, J. , Chan, C.X. , De Clerck, O. , Cock, J.M. , Coelho, S.M. , Gachon, C. , Grossman, A.R. et al. (2017) The algal revolution. Trends Plant Sci. 22, 726–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/CSP1720/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/L014130/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/R021554/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/T/000PR9819/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Research Materials
