

A primer to directed evolution: current methodologies and future directions
- PMID: 37034405
- PMCID: PMC10074555
- DOI: 10.1039/d2cb00231k
A primer to directed evolution: current methodologies and future directions
Abstract
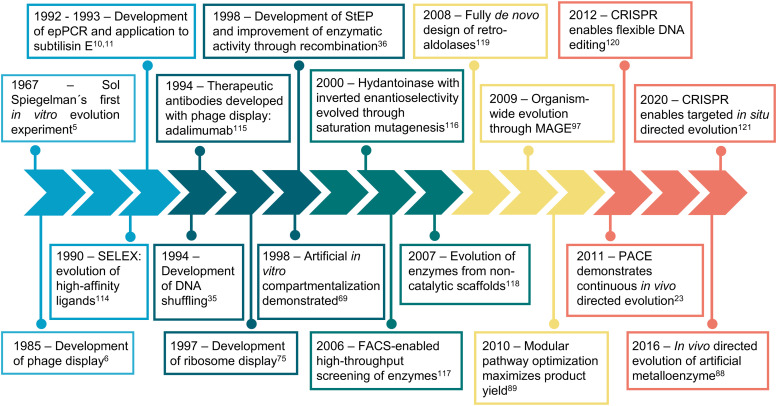
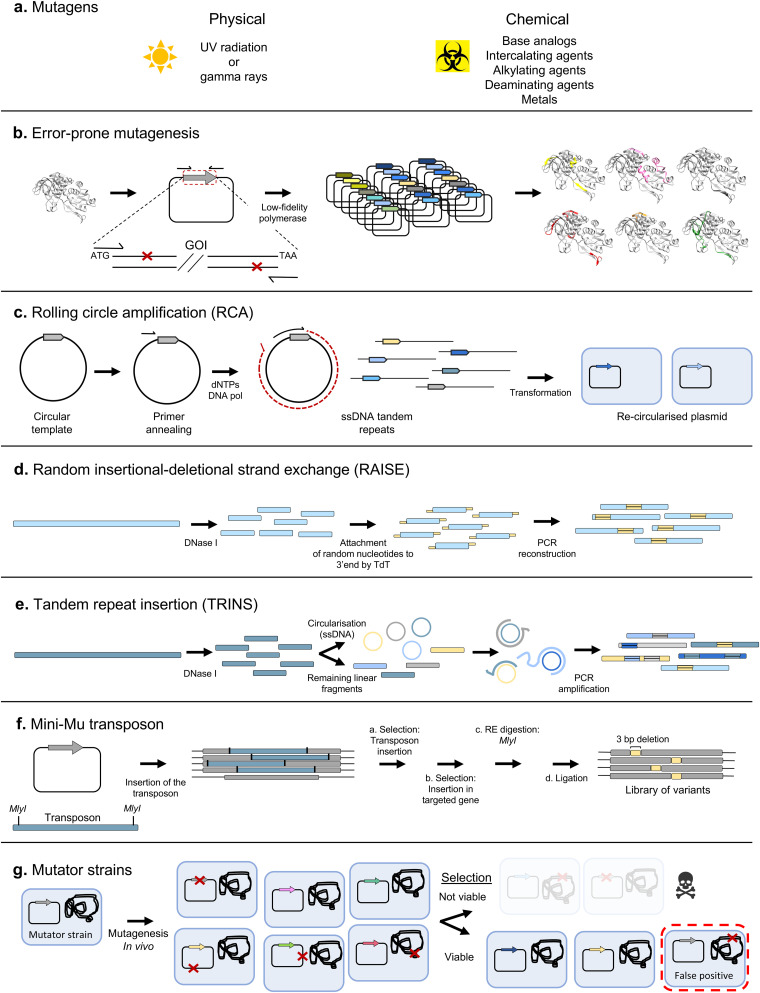
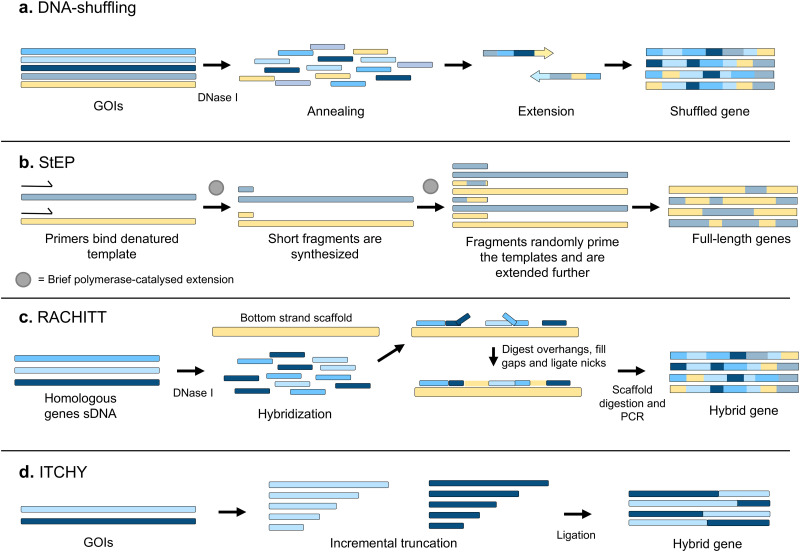
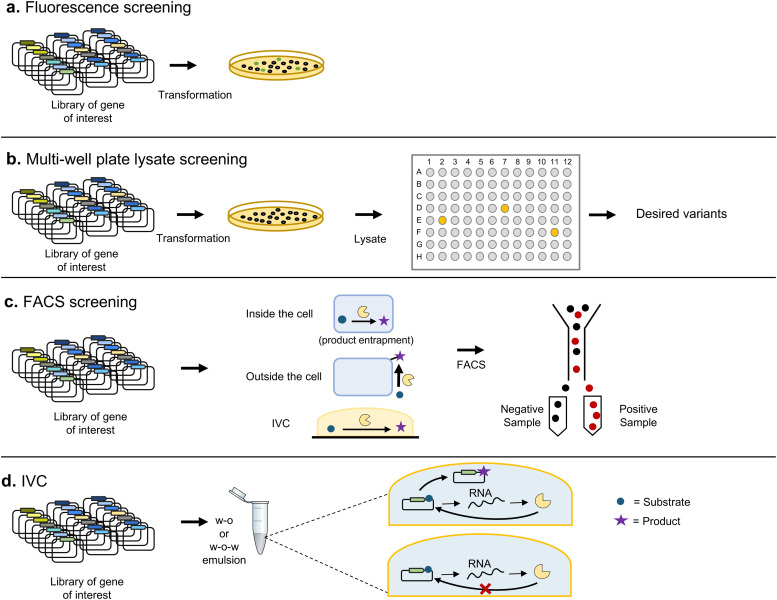
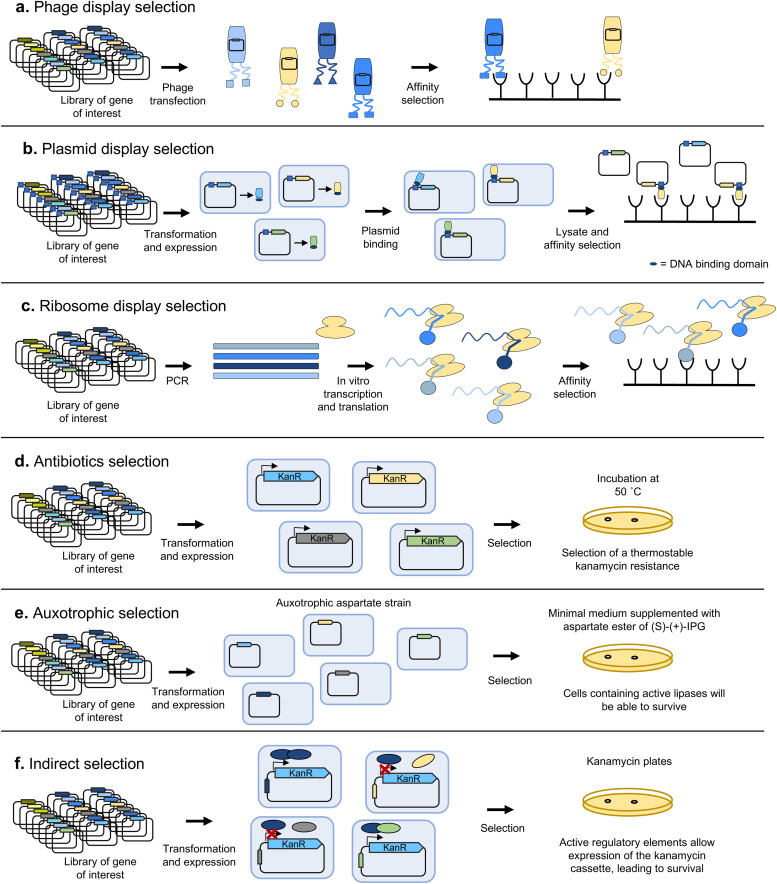
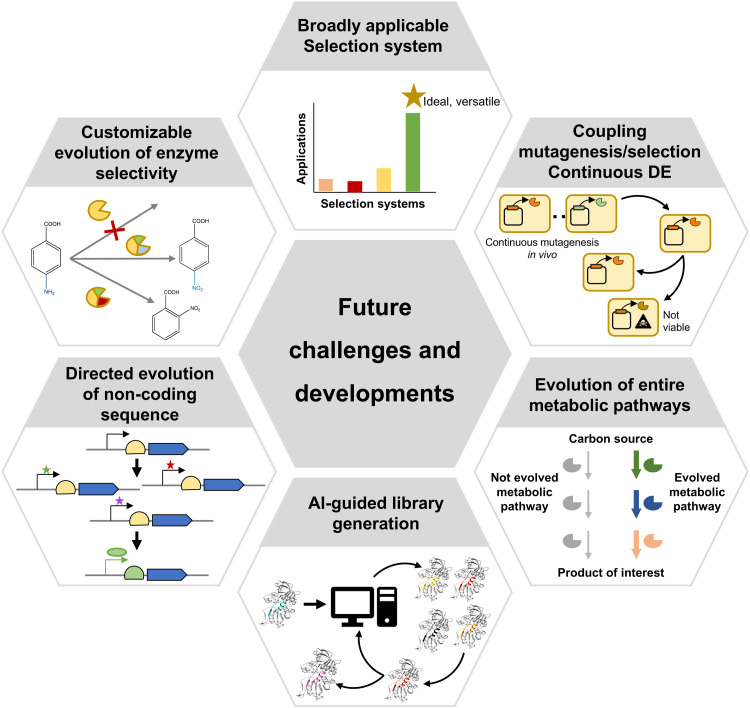
Directed evolution is one of the most powerful tools for protein engineering and functions by harnessing natural evolution, but on a shorter timescale. It enables the rapid selection of variants of biomolecules with properties that make them more suitable for specific applications. Since the first in vitro evolution experiments performed by Sol Spiegelman in 1967, a wide range of techniques have been developed to tackle the main two steps of directed evolution: genetic diversification (library generation), and isolation of the variants of interest. This review covers the main modern methodologies, discussing the advantages and drawbacks of each, and hence the considerations for designing directed evolution experiments. Furthermore, the most recent developments are discussed, showing how advances in the handling of ever larger library sizes are enabling new research questions to be tackled.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts to declare.
Figures

References
-
- Johannes T., Simurdiak M. R. and Zhao H., Biocatalysis, in Encyclopedia of Chemical Processing, CRC Press, 2005, pp. 101–110
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources