

Tom40 in cholesterol transport
- PMID: 37035007
- PMCID: PMC10074151
- DOI: 10.1016/j.isci.2023.106386
Tom40 in cholesterol transport
Abstract
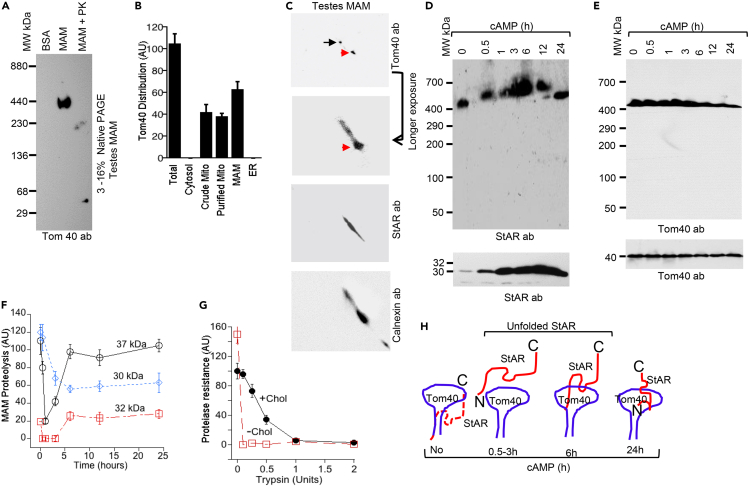
Cholesterol initiates steroid metabolism in adrenal and gonadal mitochondria, which is essential for all mammalian survival. During stress an increased cholesterol transport rapidly increases steroidogenesis; however, the mechanism of mitochondrial cholesterol transport is unknown. Using rat testicular tissue and mouse Leydig (MA-10) cells, we report for the first time that mitochondrial translocase of outer mitochondrial membrane (OMM), Tom40, is central in cholesterol transport. Cytoplasmic cholesterol-lipids complex containing StAR protein move from the mitochondria-associated ER membrane (MAM) to the OMM, increasing cholesterol load. Tom40 interacts with StAR at the OMM increasing cholesterol transport into mitochondria. An absence of Tom40 disassembles complex formation and inhibits mitochondrial cholesterol transport and steroidogenesis. Therefore, Tom40 is essential for rapid mitochondrial cholesterol transport to initiate, maintain, and regulate activity.
Keywords: Biomolecules; Cell biology; Protein folding.
© 2023 The Author(s).
Conflict of interest statement
The authors declare no competing interest.
Figures







References
-
- Clark B.J., Wells J., King S.R., Stocco D.M. The purification, cloning and expression of a novel luteinizing hormone-induced mitochondrial protein in MA-10 mouse Leydig tumor cells. Characterization of the steroidogenic acute regulatory protein (StAR) J. Biol. Chem. 1994;269:28314–28322. - PubMed
-
- Tsujishita Y., Hurley J.H. Structure and lipid transport mechanism of a StAR-related domain. Nat. Struct. Biol. 2000;7:408–414. - PubMed
-
- Artemenko I.P., Zhao D., Hales D.B., Hales K.H., Jefcoate C.R. Mitochondrial processing of newly synthesized steroidogenic acute regulatory protein (StAR), but not total StAR, mediates cholesterol transfer to cytochrome P450 side chain cleavage enzyme in adrenal cells. J. Biol. Chem. 2001;276:46583–46596. - PubMed
-
- Wang X., Liu Z., Eimerl S., Timberg R., Weiss A.M., Orly J., Stocco D.M. Effect of truncated forms of the steroidogenic acute regulatory (StAR) protein on intramitochondrial cholesterol transfer. Endocrinology. 1998;139:3903–3912. - PubMed
LinkOut - more resources
Full Text Sources
