

comCDE (Competence) Operon Is Regulated by CcpA in Streptococcus pneumoniae D39
- PMID: 37036382
- PMCID: PMC10269683
- DOI: 10.1128/spectrum.00012-23
comCDE (Competence) Operon Is Regulated by CcpA in Streptococcus pneumoniae D39
Abstract
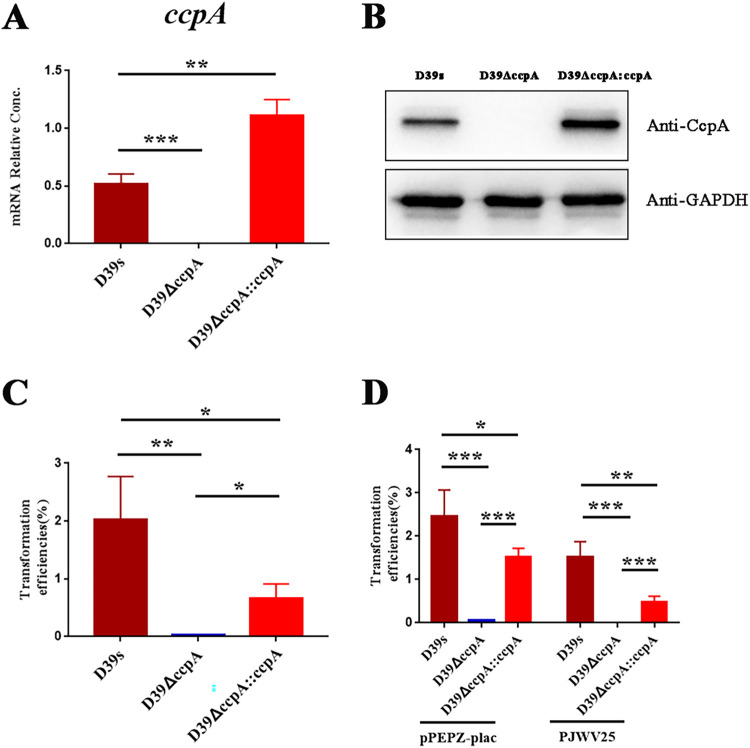
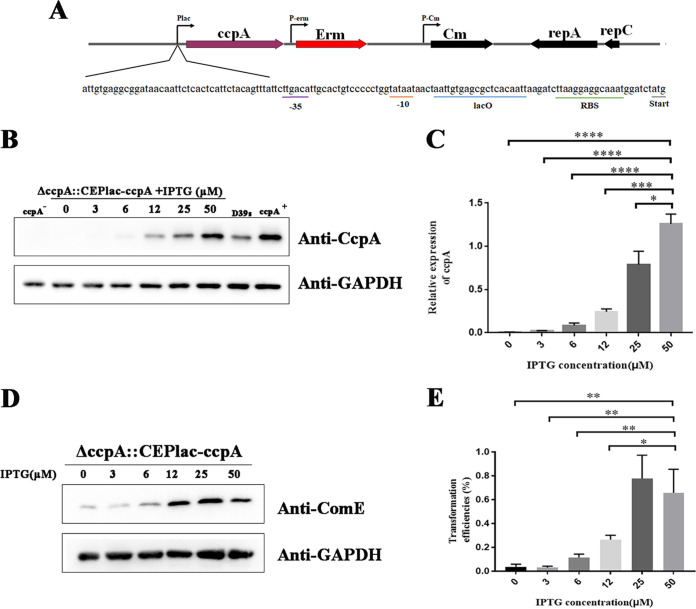
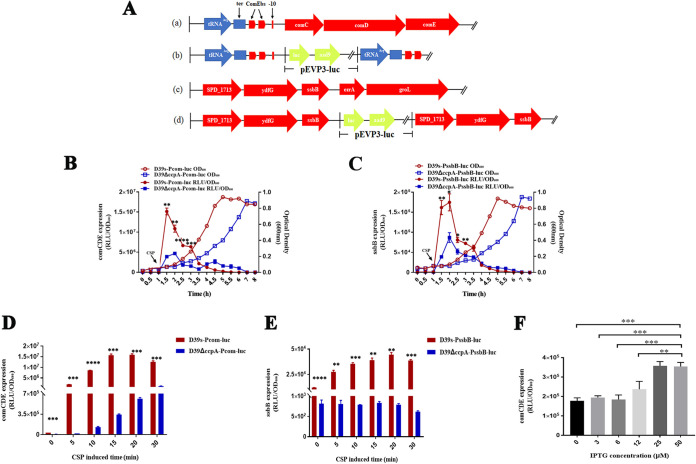
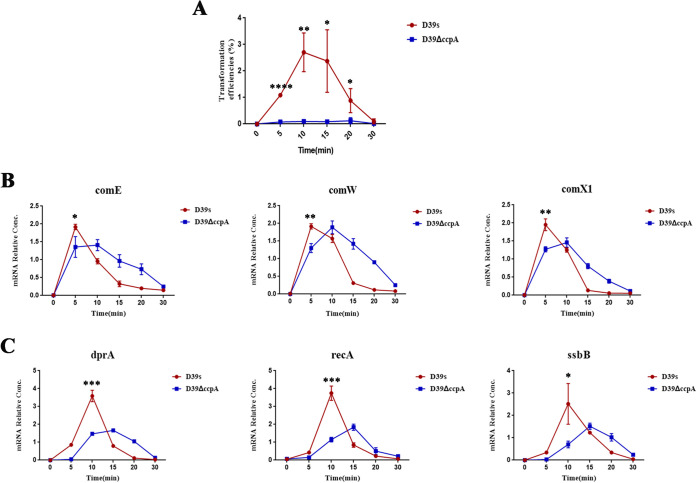
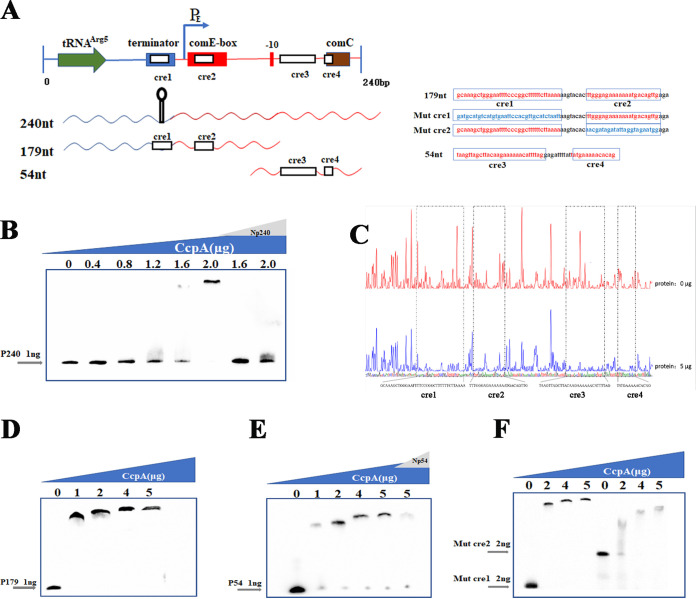
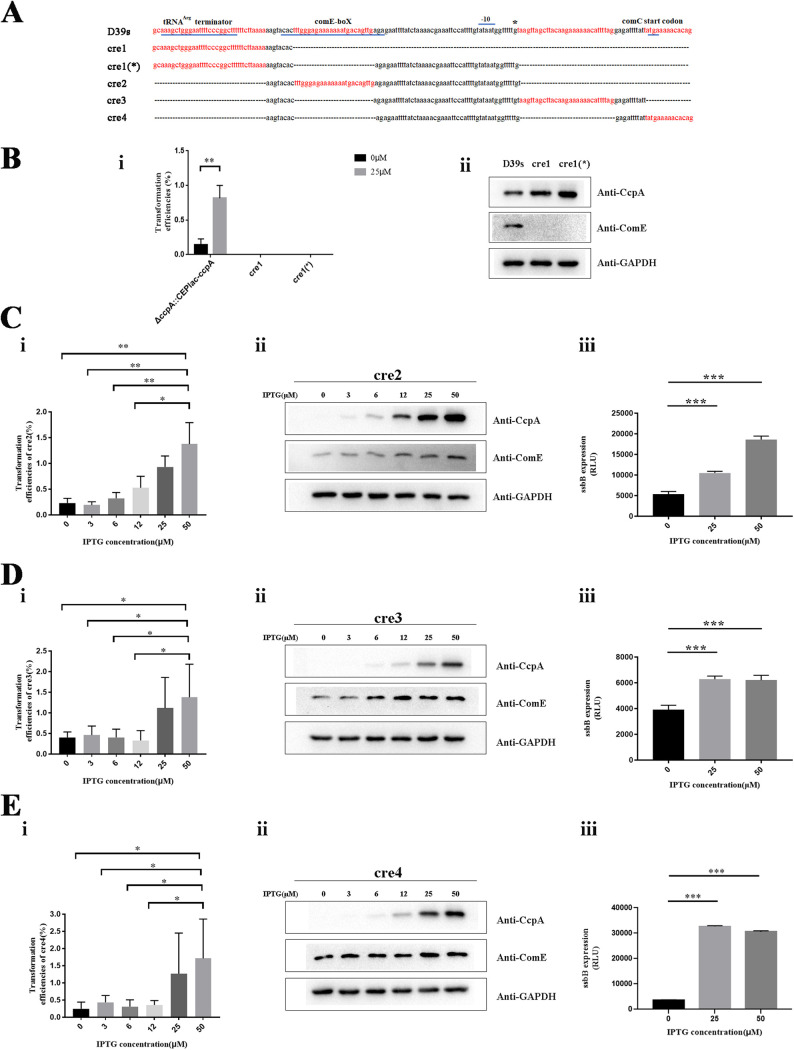
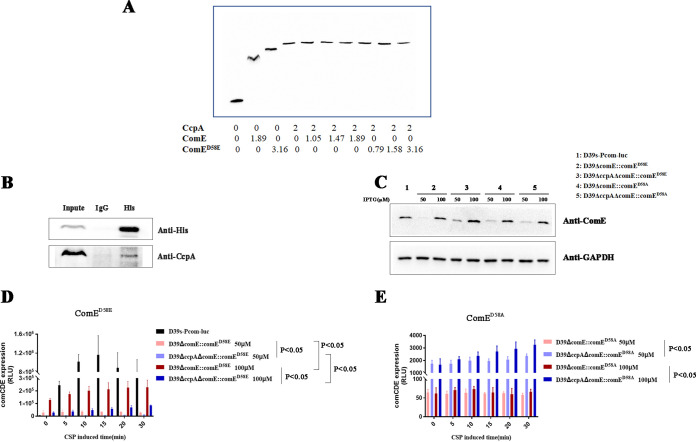
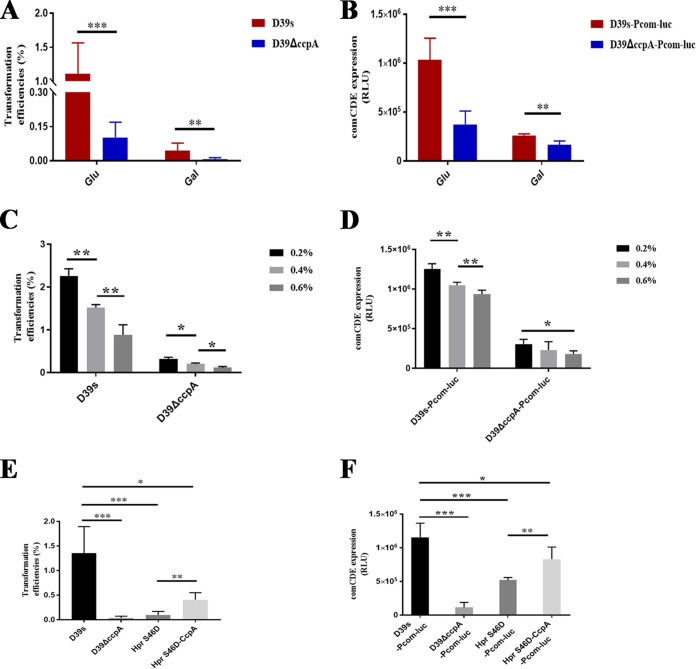
Natural transformation plays an important role in the formation of drug-resistant bacteria. Exploring the regulatory mechanism of natural transformation can aid the discovery of new antibacterial targets and reduce the emergence of drug-resistant bacteria. Competence is a prerequisite of natural transformation in Streptococcus pneumoniae, in which comCDE operon is the core regulator of competence. To date, only ComE has been shown to directly regulate comCDE transcription. In this study, a transcriptional regulator, the catabolite control protein A (CcpA), was identified that directly regulated comCDE transcription. We confirmed that CcpA binds to the cis-acting catabolite response elements (cre) in the comCDE promoter region to regulate comCDE transcription and transformation. Moreover, CcpA can coregulate comCDE transcription with phosphorylated and dephosphorylated ComE. Regulation of comCDE transcription and transformation by CcpA was also affected by carbon source signals. Together, these insights demonstrate the versatility of CcpA and provide a theoretical basis for reducing the emergence of drug-resistant bacteria. IMPORTANCE Streptococcus pneumoniae is a major cause of bacterial infections in humans, such as pneumonia, bacteremia, meningitis, otitis media, and sinusitis. Like most streptococci, S. pneumoniae is naturally competent and employs this ability to augment its adaptive evolution. The current study illustrates CcpA, a carbon catabolite regulator, can participate in the competence process by regulating comCDE transcription, and this process is regulated by different carbon source signals. These hidden abilities are likely critical for adaptation and colonization in the environment.
Keywords: CcpA; Streptococcus pneumoniae D39; competence; transformation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
The carbon catabolite repressor CcpA mediates optimal competence development in Streptococcus oligofermentans through post-transcriptional regulation.Mol Microbiol. 2019 Aug;112(2):552-568. doi: 10.1111/mmi.14274. Epub 2019 May 21. Mol Microbiol. 2019. PMID: 31074889
-
Cross-regulation of competence pheromone production and export in the early control of transformation in Streptococcus pneumoniae.Mol Microbiol. 2000 Nov;38(4):867-78. doi: 10.1046/j.1365-2958.2000.02187.x. Mol Microbiol. 2000. PMID: 11115120
-
CcpA-dependent and -independent control of beta-galactosidase expression in Streptococcus pneumoniae occurs via regulation of an upstream phosphotransferase system-encoding operon.J Bacteriol. 2007 Jul;189(14):5183-92. doi: 10.1128/JB.00449-07. Epub 2007 May 11. J Bacteriol. 2007. PMID: 17496092 Free PMC article.
-
Quorum Sensing Regulation of Competence and Bacteriocins in Streptococcus pneumoniae and mutans.Genes (Basel). 2017 Jan 5;8(1):15. doi: 10.3390/genes8010015. Genes (Basel). 2017. PMID: 28067778 Free PMC article. Review.
-
Regulation of competence for natural transformation in streptococci.Infect Genet Evol. 2015 Jul;33:343-60. doi: 10.1016/j.meegid.2014.09.010. Epub 2014 Sep 16. Infect Genet Evol. 2015. PMID: 25236918 Review.
Cited by
-
Multitasking functions of bacterial extracellular DNA in biofilms.J Bacteriol. 2024 Apr 18;206(4):e0000624. doi: 10.1128/jb.00006-24. Epub 2024 Mar 6. J Bacteriol. 2024. PMID: 38445859 Free PMC article. Review.
-
Unveiling the regulatory network controlling natural transformation in lactococci.PLoS Genet. 2024 Jul 1;20(7):e1011340. doi: 10.1371/journal.pgen.1011340. eCollection 2024 Jul. PLoS Genet. 2024. PMID: 38950059 Free PMC article.
-
Transcriptional regulation of TacL-mediated lipoteichoic acids biosynthesis by ComE during competence impacts pneumococcal transformation.Front Cell Infect Microbiol. 2024 May 8;14:1375312. doi: 10.3389/fcimb.2024.1375312. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38779562 Free PMC article.
-
Microbial Community Changes in Silkworms Suspected of Septicemia and Identification of Serratia sp.Int J Mol Sci. 2024 Apr 2;25(7):3957. doi: 10.3390/ijms25073957. Int J Mol Sci. 2024. PMID: 38612767 Free PMC article.
-
The global proteome of Streptococcus pneumoniae EF3030 under nutrient-defined in vitro conditions.Front Cell Infect Microbiol. 2025 Jul 11;15:1606161. doi: 10.3389/fcimb.2025.1606161. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 40718681 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
