

An ancient metalloenzyme evolves through metal preference modulation
- PMID: 37037909
- PMCID: PMC10172142
- DOI: 10.1038/s41559-023-02012-0
An ancient metalloenzyme evolves through metal preference modulation
Abstract
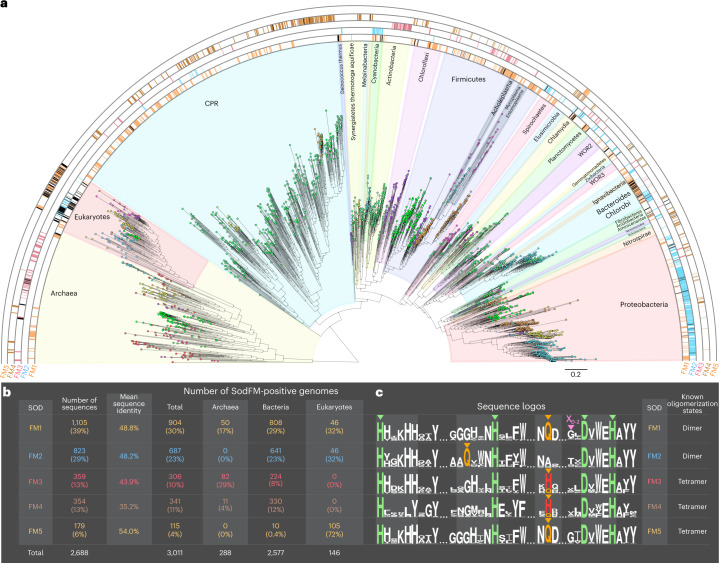
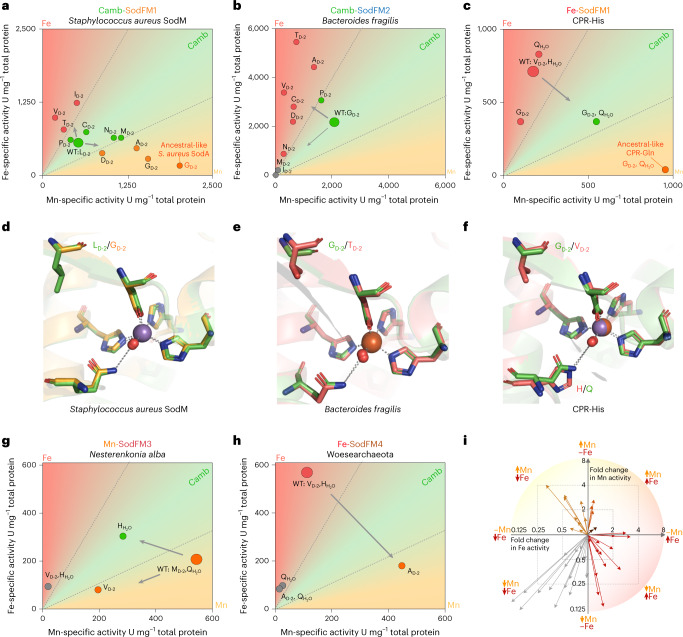
Evolution creates functional diversity of proteins, the essential building blocks of all biological systems. However, studies of natural proteins sampled across the tree of life and evaluated in a single experimental system are lacking. Almost half of enzymes require metals, and metalloproteins tend to optimally utilize the physicochemical properties of a specific metal co-factor. Life must adapt to changes in metal bioavailability, including those during the transition from anoxic to oxic Earth or pathogens' exposure to nutritional immunity. These changes can challenge the ability of metalloenzymes to maintain activity, presumptively driving their evolution. Here we studied metal-preference evolution within the natural diversity of the iron/manganese superoxide dismutase (SodFM) family of reactive oxygen species scavengers. We identified and experimentally verified residues with conserved roles in determining metal preference that, when combined with an understanding of the protein's evolutionary history, improved prediction of metal utilization across the five SodFM subfamilies defined herein. By combining phylogenetics, biochemistry and structural biology, we demonstrate that SodFM metal utilization can be evolutionarily fine tuned by sliding along a scale between perfect manganese and iron specificities. Over the history of life, SodFM metal preference has been modulated multiple independent times within different evolutionary and ecological contexts, and can be changed within short evolutionary timeframes.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures












References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
