

Intestinal α-Defensins Play a Minor Role in Modulating the Small Intestinal Microbiota Composition as Compared to Diet
- PMID: 37039638
- PMCID: PMC10269482
- DOI: 10.1128/spectrum.00567-23
Intestinal α-Defensins Play a Minor Role in Modulating the Small Intestinal Microbiota Composition as Compared to Diet
Abstract
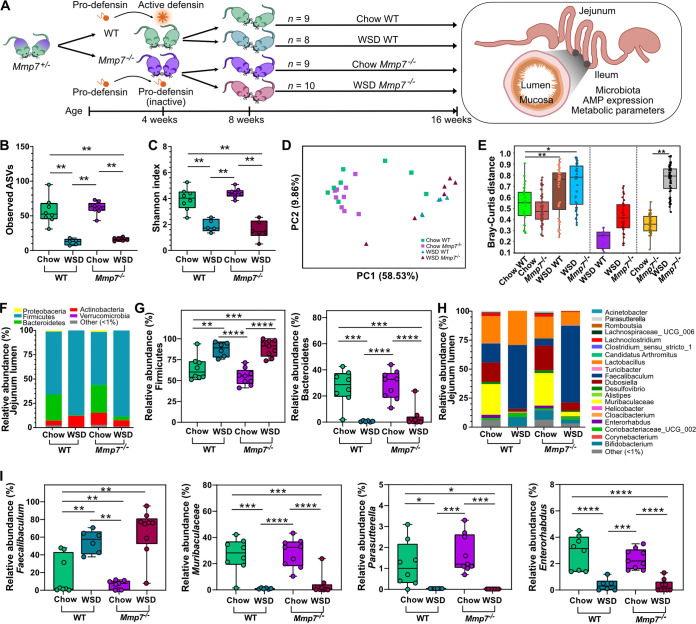
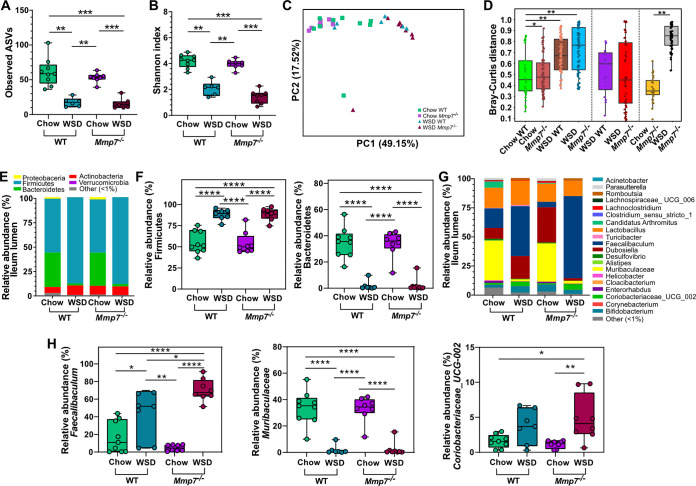
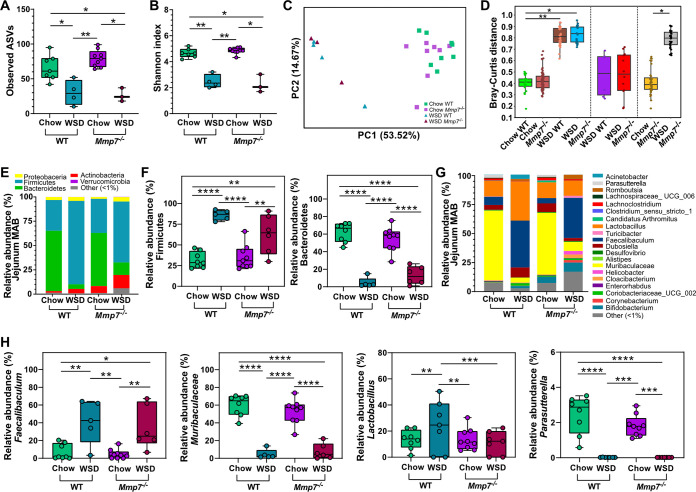
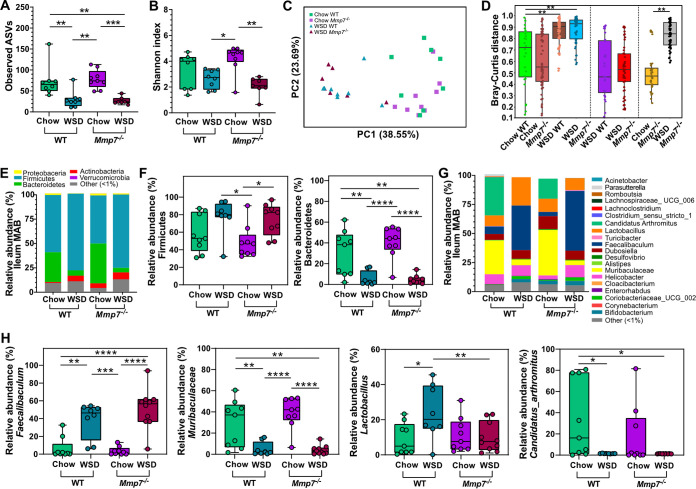
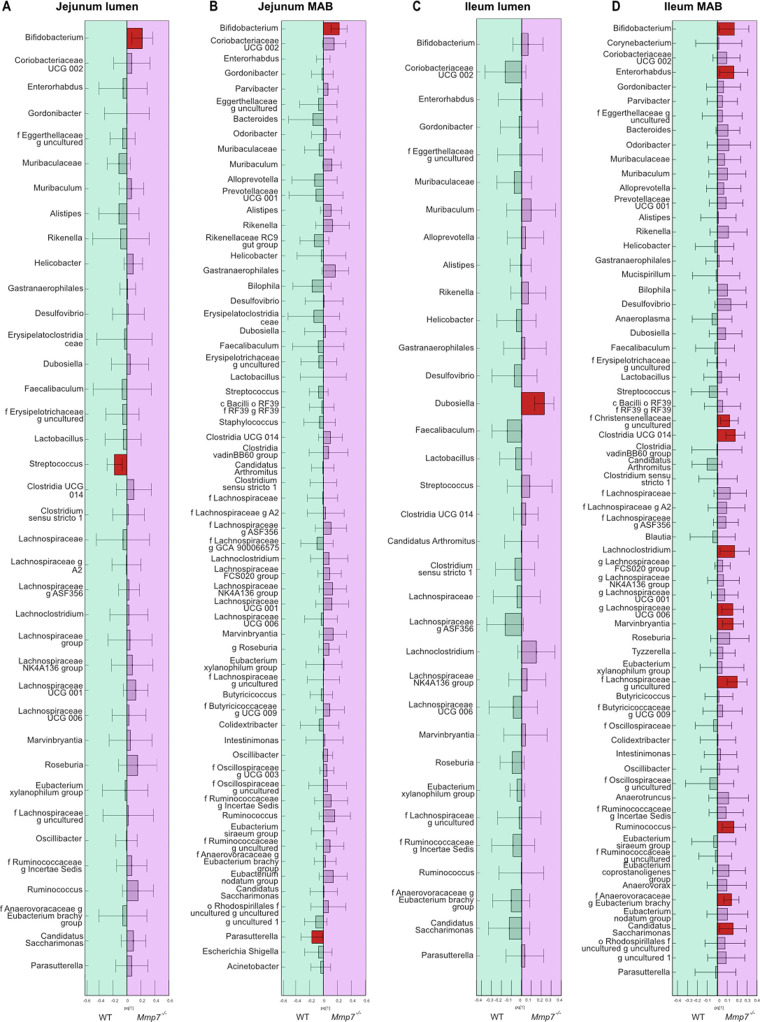
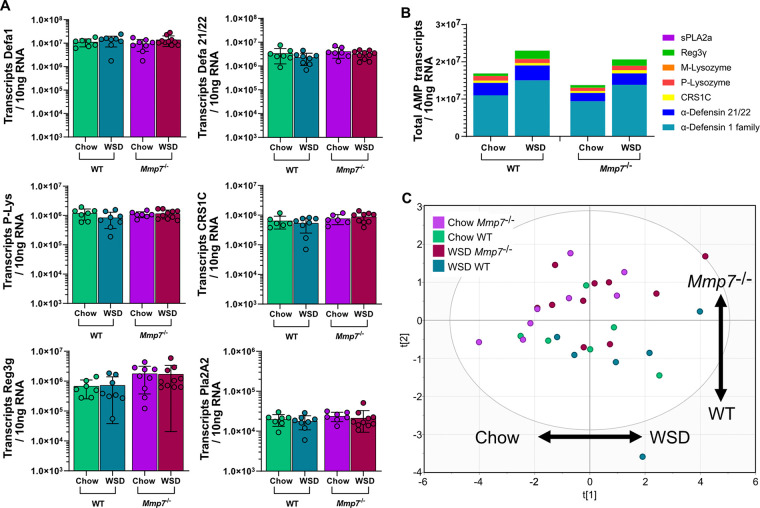
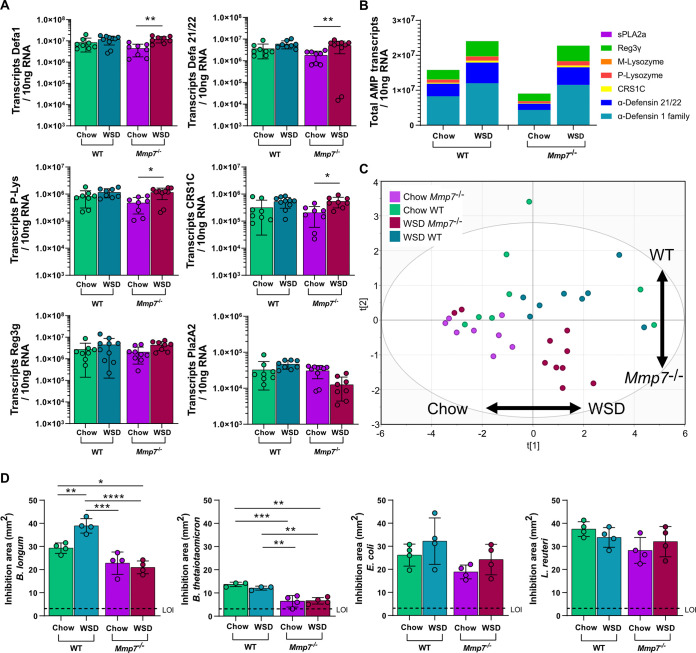
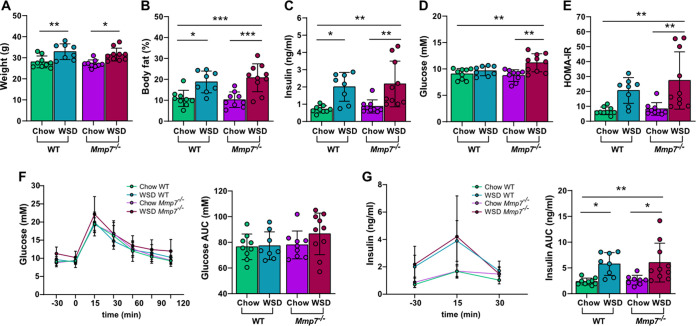
The intestinal microbiota is at the interface between the host and its environment and thus under constant exposure to host-derived and external modulators. While diet is considered to be an important external factor modulating microbiota composition, intestinal defensins, one of the major classes of antimicrobial peptides, have been described as key host effectors that shape the gut microbial community. However, since dietary compounds can affect defensin expression, thereby indirectly modulating the intestinal microbiota, their individual contribution to shaping gut microbiota composition remains to be defined. To disentangle the complex interaction among diet, defensins, and small-intestinal microbiota, we fed wild-type (WT) mice and mice lacking functionally active α-defensins (Mmp7-/- mice) either a control diet or a Western-style diet (WSD) that is rich in saturated fat and simple carbohydrates but low in dietary fiber. 16S rDNA sequencing and robust statistical analyses identified that bacterial composition was strongly affected by diet while defensins had only a minor impact. These findings were independent of sample location, with consistent results between the lumen and mucosa of the jejunum and ileum, in both mouse genotypes. However, distinct microbial taxa were also modulated by α-defensins, which was supported by differential antimicrobial activity of ileal protein extracts. As the combination of WSD and defensin deficiency exacerbated glucose metabolism, we conclude that defensins only have a fine-tuning role in shaping the small-intestinal bacterial composition and might instead be important in protecting the host against the development of diet-induced metabolic dysfunction. IMPORTANCE Alterations in the gut microbial community composition are associated with many diseases, and therefore identifying factors that shape the microbial community under homeostatic and diseased conditions may contribute to the development of strategies to correct a dysbiotic microbiota. Here, we demonstrate that a Western-style diet, as an extrinsic parameter, had a stronger impact on shaping the small intestinal bacterial composition than intestinal defensins, as an intrinsic parameter. While defensins have been previously shown to modulate bacterial composition in young mice, our study supplements these findings by showing that defensins may be less important in adult mice that harbor a mature microbial community. Nevertheless, we observed that defensins did affect the abundance of distinct bacterial taxa in adult mice and protected the host from aggravated diet-induced glucose impairments. Consequently, our study uncovers a new angle on the role of intestinal defensins in the development of metabolic diseases in adult mice.
Keywords: Western diet; antimicrobial peptides; defensins; gut microbiota; metabolic disease; mucosal barrier.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Intestinal Barrier Function and the Gut Microbiome Are Differentially Affected in Mice Fed a Western-Style Diet or Drinking Water Supplemented with Fructose.J Nutr. 2017 May;147(5):770-780. doi: 10.3945/jn.116.242859. Epub 2017 Mar 29. J Nutr. 2017. PMID: 28356436
-
Does an Apple a Day Also Keep the Microbes Away? The Interplay Between Diet, Microbiota, and Host Defense Peptides at the Intestinal Mucosal Barrier.Front Immunol. 2020 Jun 9;11:1164. doi: 10.3389/fimmu.2020.01164. eCollection 2020. Front Immunol. 2020. PMID: 32655555 Free PMC article. Review.
-
Paneth cells, defensins, and the commensal microbiota: a hypothesis on intimate interplay at the intestinal mucosa.Semin Immunol. 2007 Apr;19(2):70-83. doi: 10.1016/j.smim.2007.04.002. Epub 2007 May 7. Semin Immunol. 2007. PMID: 17485224 Review.
-
Alterations of the Viable Ileal Microbiota of the Gut Mucosa-Lymph Node Axis in Pigs Fed Phytase and Lactic Acid-Treated Cereals.Appl Environ Microbiol. 2020 Feb 3;86(4):e02128-19. doi: 10.1128/AEM.02128-19. Print 2020 Feb 3. Appl Environ Microbiol. 2020. PMID: 31757823 Free PMC article.
-
Paneth cell α-defensins HD-5 and HD-6 display differential degradation into active antimicrobial fragments.Proc Natl Acad Sci U S A. 2019 Feb 26;116(9):3746-3751. doi: 10.1073/pnas.1817376116. Epub 2019 Feb 11. Proc Natl Acad Sci U S A. 2019. PMID: 30808760 Free PMC article.
Cited by
-
Disentangling the impact of obesity, diet, host factors, and microbiota on small intestinal antimicrobial peptide expression.Gut Microbes. 2025 Dec;17(1):2536095. doi: 10.1080/19490976.2025.2536095. Epub 2025 Aug 4. Gut Microbes. 2025. PMID: 40760765 Free PMC article.
-
Defensins: Exploring Their Opposing Roles in Colorectal Cancer Progression.Cancers (Basel). 2024 Jul 23;16(15):2622. doi: 10.3390/cancers16152622. Cancers (Basel). 2024. PMID: 39123348 Free PMC article. Review.
-
Parenteral nutrition results in peripheral ileal to hepatic circadian discordance in mice.Am J Physiol Gastrointest Liver Physiol. 2024 Dec 1;327(6):G754-G764. doi: 10.1152/ajpgi.00042.2024. Epub 2024 Sep 20. Am J Physiol Gastrointest Liver Physiol. 2024. PMID: 39301965
-
Gut microbiota development across the lifespan: Disease links and health-promoting interventions.J Intern Med. 2025 Jun;297(6):560-583. doi: 10.1111/joim.20089. Epub 2025 Apr 24. J Intern Med. 2025. PMID: 40270478 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
