

The RhoGEF protein Plekhg5 regulates medioapical and junctional actomyosin dynamics of apical constriction during Xenopus gastrulation
- PMID: 37043306
- PMCID: PMC10295481
- DOI: 10.1091/mbc.E22-09-0411
The RhoGEF protein Plekhg5 regulates medioapical and junctional actomyosin dynamics of apical constriction during Xenopus gastrulation
Abstract
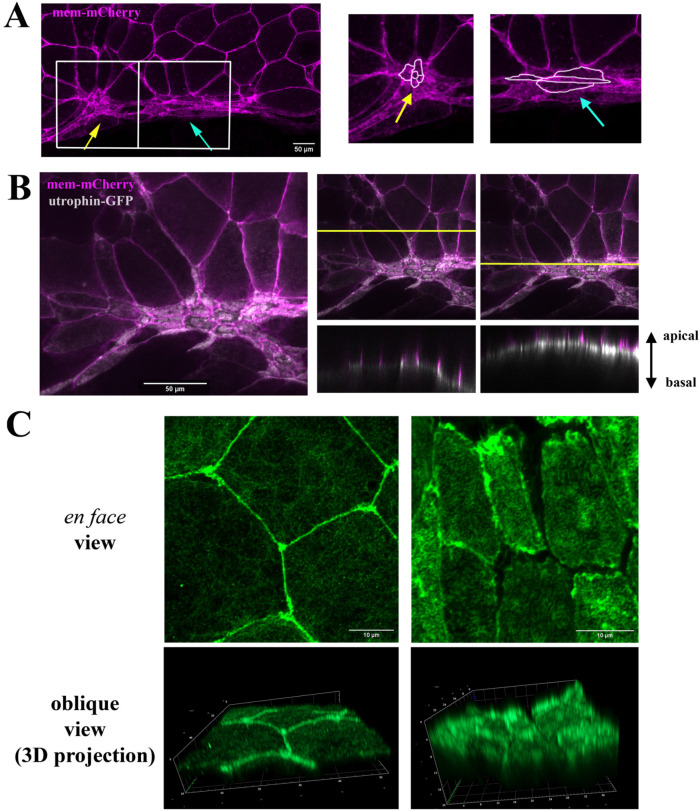
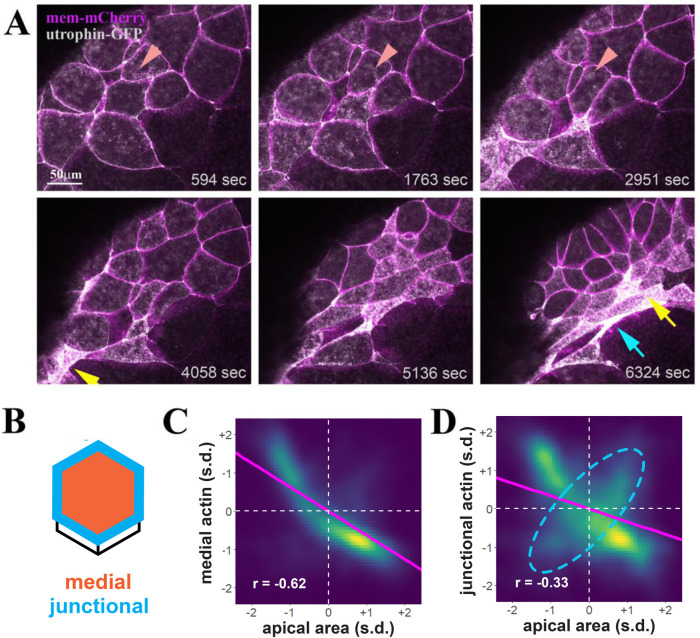
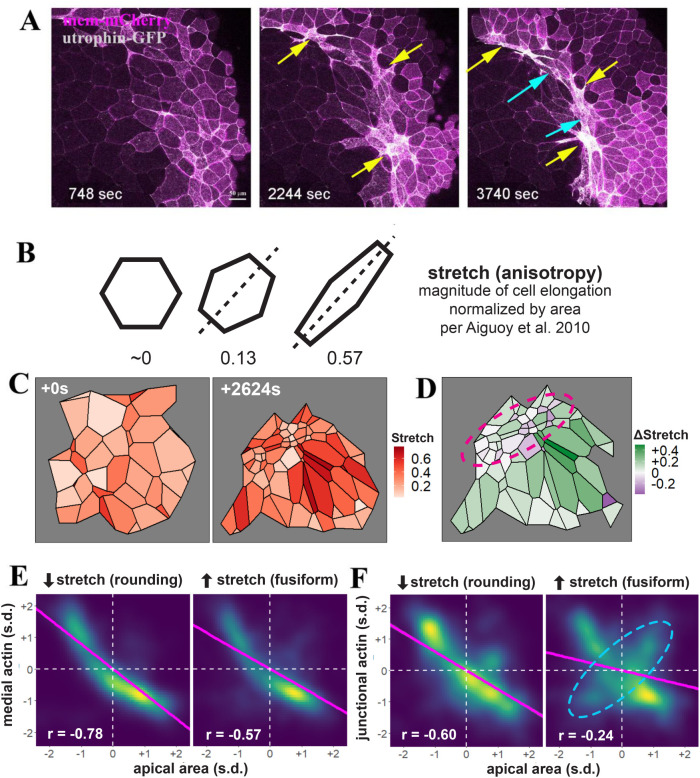
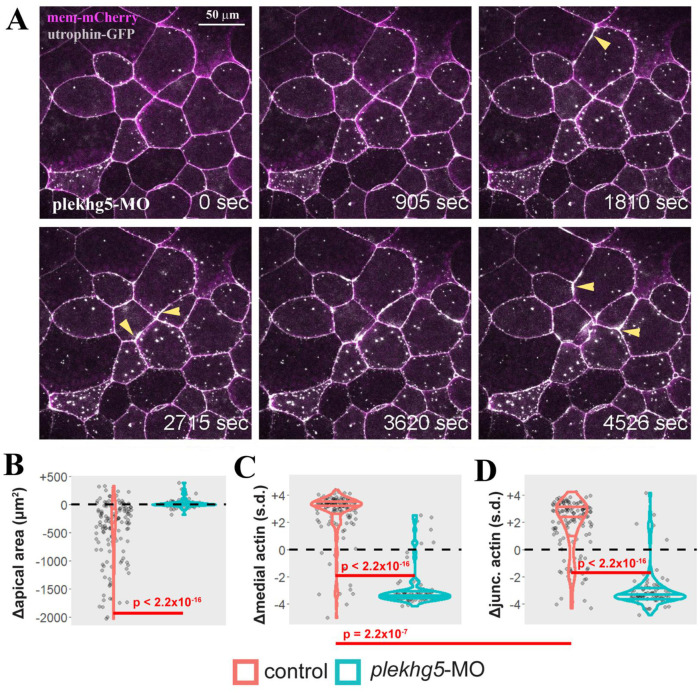
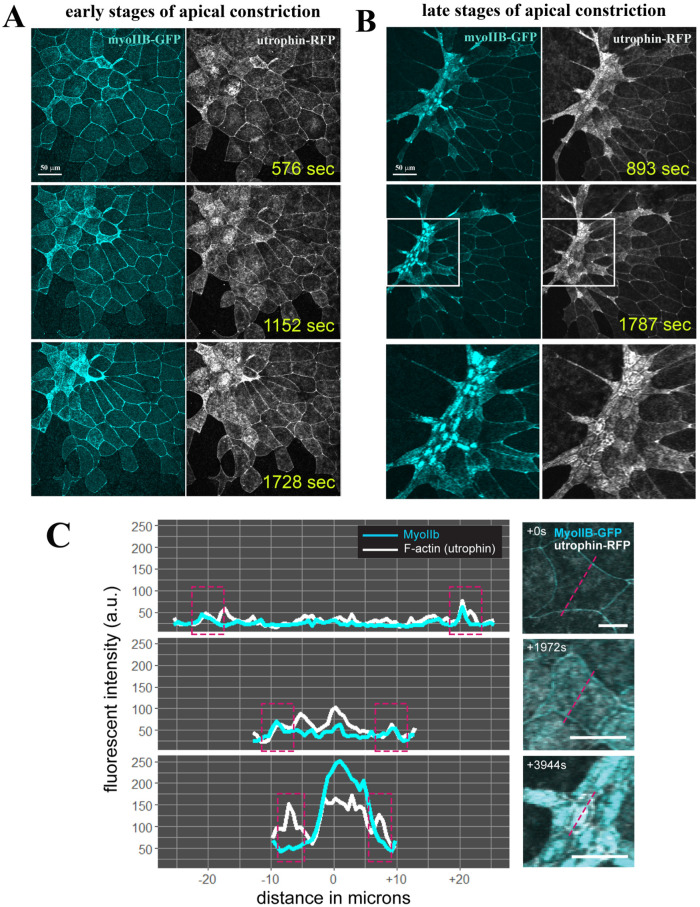
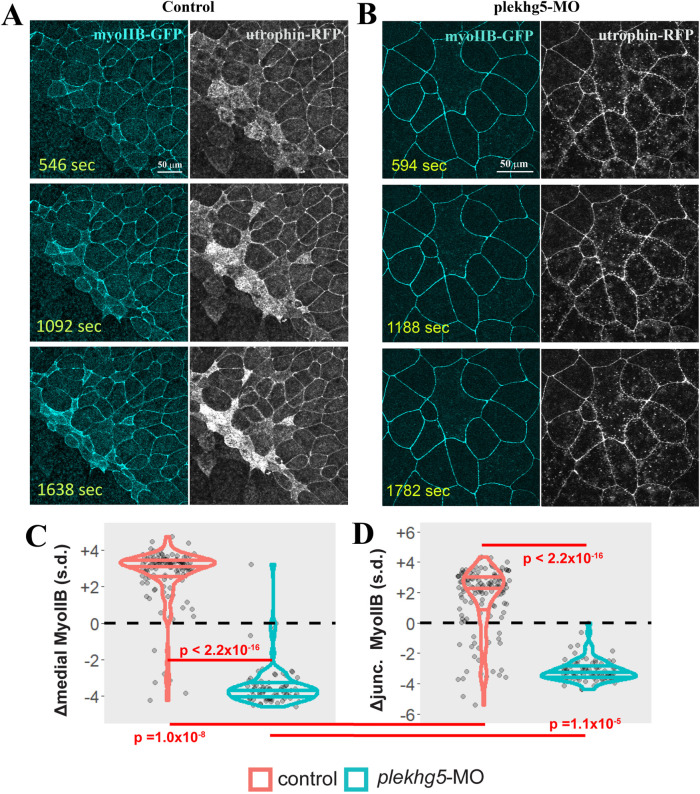
Apical constriction results in apical surface reduction in epithelial cells and is a widely used mechanism for epithelial morphogenesis. Both medioapical and junctional actomyosin remodeling are involved in apical constriction, but the deployment of medial versus junctional actomyosin and their genetic regulation in vertebrate embryonic development have not been fully described. In this study, we investigate actomyosin dynamics and their regulation by the RhoGEF protein Plekhg5 in Xenopus bottle cells. Using live imaging and quantitative image analysis, we show that bottle cells assume different shapes, with rounding bottle cells constricting earlier in small clusters followed by fusiform bottle cells forming between the clusters. Both medioapical and junctional actomyosin signals increase as surface area decreases, though correlation of apical constriction with medioapical actomyosin localization appears to be stronger. F-actin bundles perpendicular to the apical surface form in constricted cells, which may correspond to microvilli previously observed in the apical membrane. Knockdown of plekhg5 disrupts medioapical and junctional actomyosin activity and apical constriction but does not affect initial F-actin dynamics. Taking the results together, our study reveals distinct cell morphologies, uncovers actomyosin behaviors, and demonstrates the crucial role of a RhoGEF protein in controlling actomyosin dynamics during apical constriction of bottle cells in Xenopus gastrulation.
Figures
Similar articles
-
The RhoGEF protein Plekhg5 self-associates via its PH domain to regulate apical cell constriction.Mol Biol Cell. 2024 Oct 1;35(10):ar134. doi: 10.1091/mbc.E24-04-0179. Epub 2024 Aug 28. Mol Biol Cell. 2024. PMID: 39196644 Free PMC article.
-
The RhoGEF protein Plekhg5 regulates apical constriction of bottle cells during gastrulation.Development. 2018 Dec 12;145(24):dev168922. doi: 10.1242/dev.168922. Development. 2018. PMID: 30446627 Free PMC article.
-
Actomyosin contractility and microtubules drive apical constriction in Xenopus bottle cells.Dev Biol. 2007 Nov 1;311(1):40-52. doi: 10.1016/j.ydbio.2007.08.010. Epub 2007 Aug 10. Dev Biol. 2007. PMID: 17868669 Free PMC article.
-
Uncorking gastrulation: the morphogenetic movement of bottle cells.Wiley Interdiscip Rev Dev Biol. 2012 Mar-Apr;1(2):286-93. doi: 10.1002/wdev.19. Epub 2011 Dec 12. Wiley Interdiscip Rev Dev Biol. 2012. PMID: 23801442 Free PMC article. Review.
-
Apical constriction in morphogenesis: From actomyosin architecture to regulatory networks.Curr Opin Cell Biol. 2025 Aug;95:102562. doi: 10.1016/j.ceb.2025.102562. Epub 2025 Jun 12. Curr Opin Cell Biol. 2025. PMID: 40513206 Review.
Cited by
-
Mechanical cues polarize ADIP protein complexes to control vertebrate morphogenesis and wound healing.Curr Biol. 2025 Jul 21;35(14):3315-3326.e4. doi: 10.1016/j.cub.2025.05.069. Epub 2025 Jun 24. Curr Biol. 2025. PMID: 40562038
-
Architecture of the cortical actomyosin network driving apical constriction in C. elegans.J Cell Biol. 2023 Sep 4;222(9):e202302102. doi: 10.1083/jcb.202302102. Epub 2023 Jun 23. J Cell Biol. 2023. PMID: 37351566 Free PMC article.
-
Architecture of the cortical actomyosin network driving apical constriction in C. elegans.bioRxiv [Preprint]. 2023 Feb 1:2023.01.30.526280. doi: 10.1101/2023.01.30.526280. bioRxiv. 2023. Update in: J Cell Biol. 2023 Sep 4;222(9):e202302102. doi: 10.1083/jcb.202302102. PMID: 36778218 Free PMC article. Updated. Preprint.
-
The RhoGEF protein Plekhg5 self-associates via its PH domain to regulate apical cell constriction.Mol Biol Cell. 2024 Oct 1;35(10):ar134. doi: 10.1091/mbc.E24-04-0179. Epub 2024 Aug 28. Mol Biol Cell. 2024. PMID: 39196644 Free PMC article.
-
Mechanical control of neural plate folding by apical domain alteration.Nat Commun. 2023 Dec 20;14(1):8475. doi: 10.1038/s41467-023-43973-x. Nat Commun. 2023. PMID: 38123550 Free PMC article.
References
-
- Aigouy B, Farhadifar R, Staple DB, Sagner A, Roper JC, Julicher F, Eaton S (2010). Cell flow reorients the axis of planar polarity in the wing epithelium of Drosophila. Cell 142, 773–786. - PubMed
-
- Aigouy B, Umetsu D, Eaton S (2016). Segmentation and quantitative analysis of epithelial tissues. Methods Mol Biol 1478, 227–239. - PubMed
-
- An Y, Xue G, Shaobo Y, Mingxi D, Zhou X, Yu W, Ishibashi T, Zhang L, Yan Y (2017). Apical constriction is driven by a pulsatile apical myosin network in delaminating Drosophila neuroblasts. Development 144, 2153–2164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
