

Cation leak through the ATP1A3 pump causes spasticity and intellectual disability
- PMID: 37043503
- PMCID: PMC10393399
- DOI: 10.1093/brain/awad124
Cation leak through the ATP1A3 pump causes spasticity and intellectual disability
Abstract
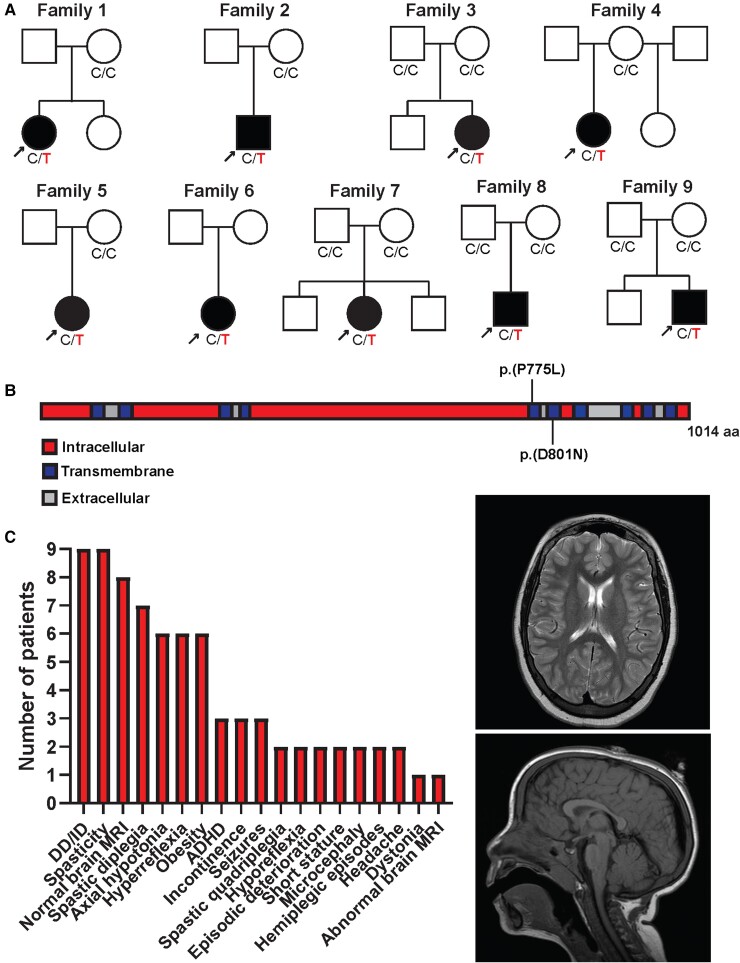
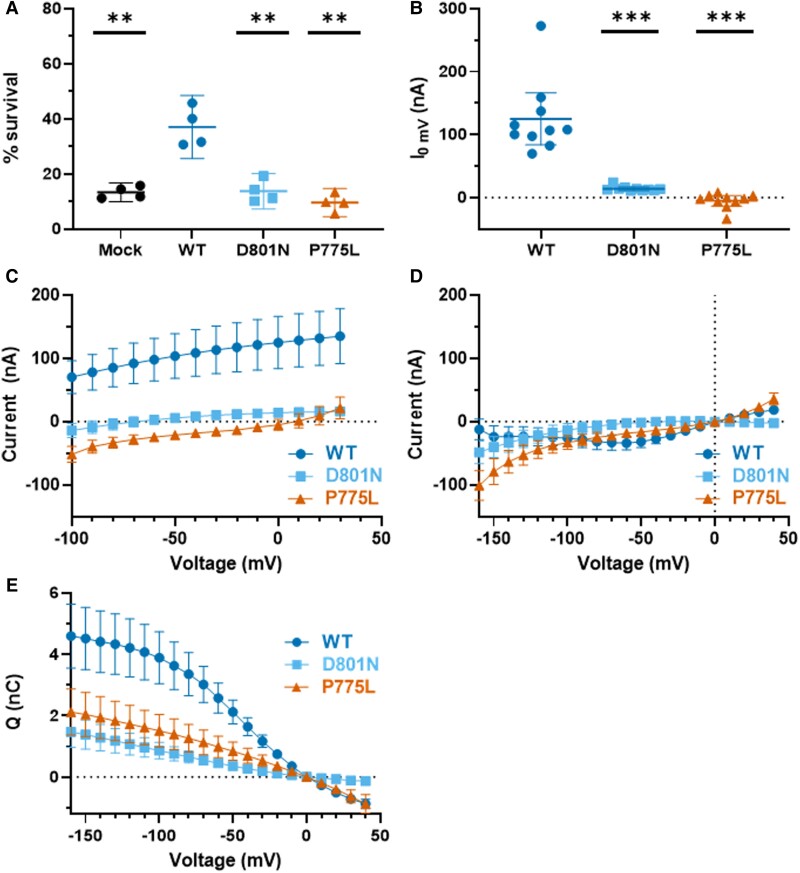
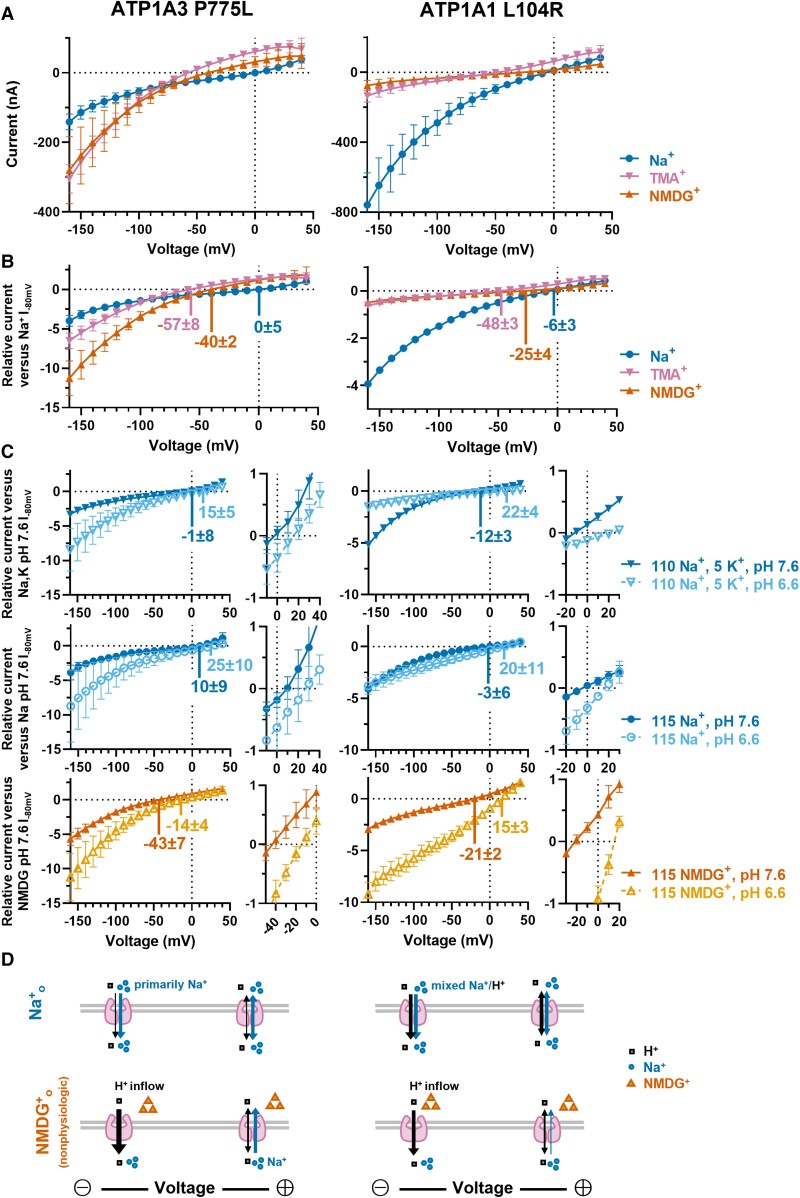
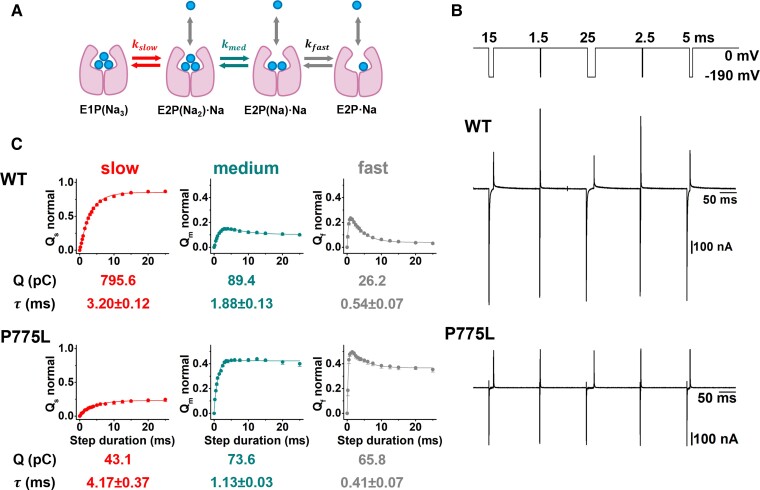
ATP1A3 encodes the α3 subunit of the sodium-potassium ATPase, one of two isoforms responsible for powering electrochemical gradients in neurons. Heterozygous pathogenic ATP1A3 variants produce several distinct neurological syndromes, yet the molecular basis for phenotypic variability is unclear. We report a novel recurrent variant, ATP1A3(NM_152296.5):c.2324C>T; p.(Pro775Leu), in nine individuals associated with the primary clinical features of progressive or non-progressive spasticity and developmental delay/intellectual disability. No patients fulfil diagnostic criteria for ATP1A3-associated syndromes, including alternating hemiplegia of childhood, rapid-onset dystonia-parkinsonism or cerebellar ataxia-areflexia-pes cavus-optic atrophy-sensorineural hearing loss (CAPOS), and none were suspected of having an ATP1A3-related disorder. Uniquely among known ATP1A3 variants, P775L causes leakage of sodium ions and protons into the cell, associated with impaired sodium binding/occlusion kinetics favouring states with fewer bound ions. These phenotypic and electrophysiologic studies demonstrate that ATP1A3:c.2324C>T; p.(Pro775Leu) results in mild ATP1A3-related phenotypes resembling complex hereditary spastic paraplegia or idiopathic spastic cerebral palsy. Cation leak provides a molecular explanation for this genotype-phenotype correlation, adding another mechanism to further explain phenotypic variability and highlighting the importance of biophysical properties beyond ion transport rate in ion transport diseases.
Keywords: ATP1A3; neurodevelopmental disorders; sodium-potassium ATPase; spastic paraparesis; spasticity.
Published by Oxford University Press on behalf of the Guarantors of Brain 2023.
Conflict of interest statement
The authors report no competing interests.
Figures
References
-
- de Carvalho Aguiar P, Sweadner KJ, Penniston JT, et al. . Mutations in the Na+/K+-ATPase alpha3 gene ATP1A3 are associated with rapid-onset dystonia parkinsonism. Neuron. 2004;43:169–175. - PubMed
-
- Rosewich H, Thiele H, Ohlenbusch A, et al. . Heterozygous de-novo mutations in ATP1A3 in patients with alternating hemiplegia of childhood: a whole-exome sequencing gene-identification study. Lancet Neurol. 2012;11:764–773. - PubMed
