

The Origin of Floral Quartet Formation-Ancient Exon Duplications Shaped the Evolution of MIKC-type MADS-domain Transcription Factor Interactions
- PMID: 37043523
- PMCID: PMC10152394
- DOI: 10.1093/molbev/msad088
The Origin of Floral Quartet Formation-Ancient Exon Duplications Shaped the Evolution of MIKC-type MADS-domain Transcription Factor Interactions
Abstract
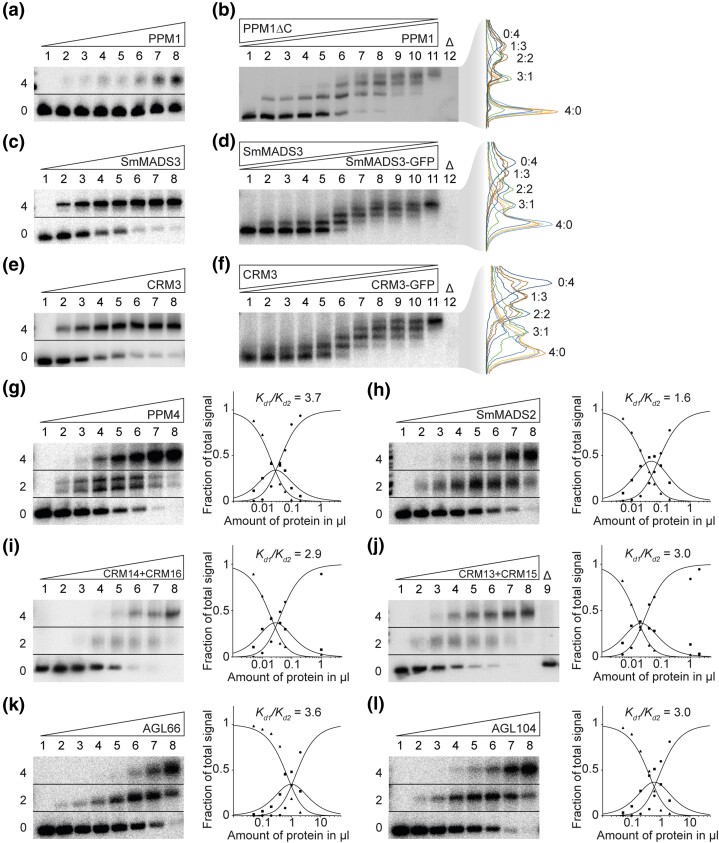
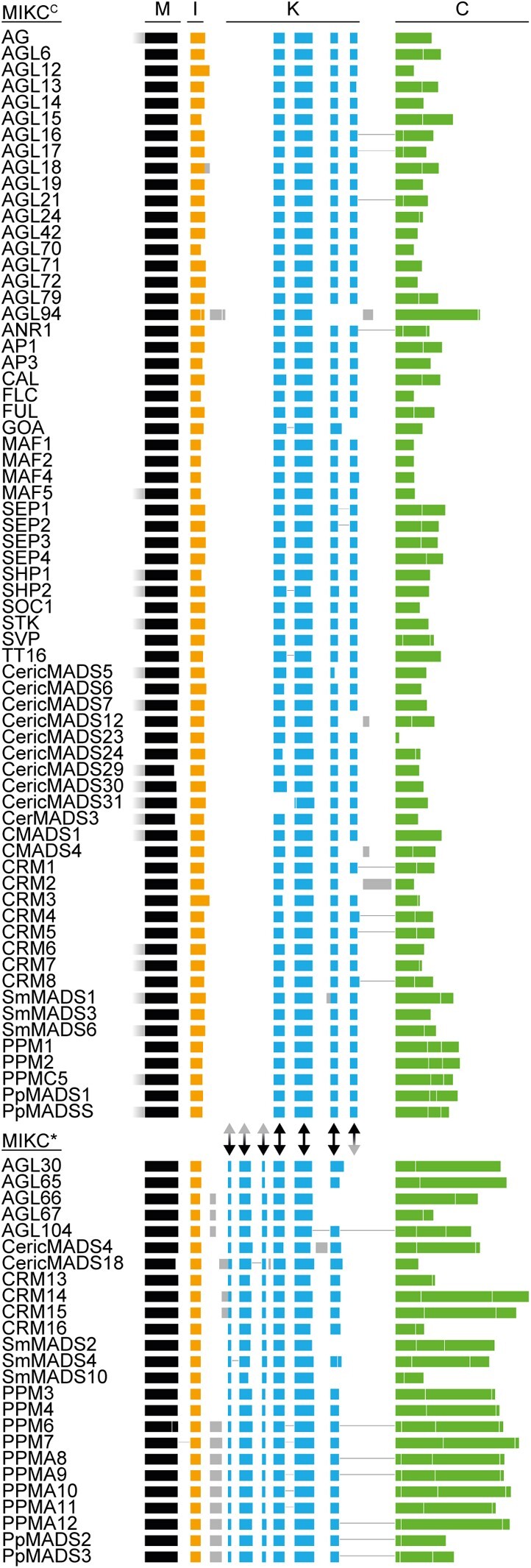
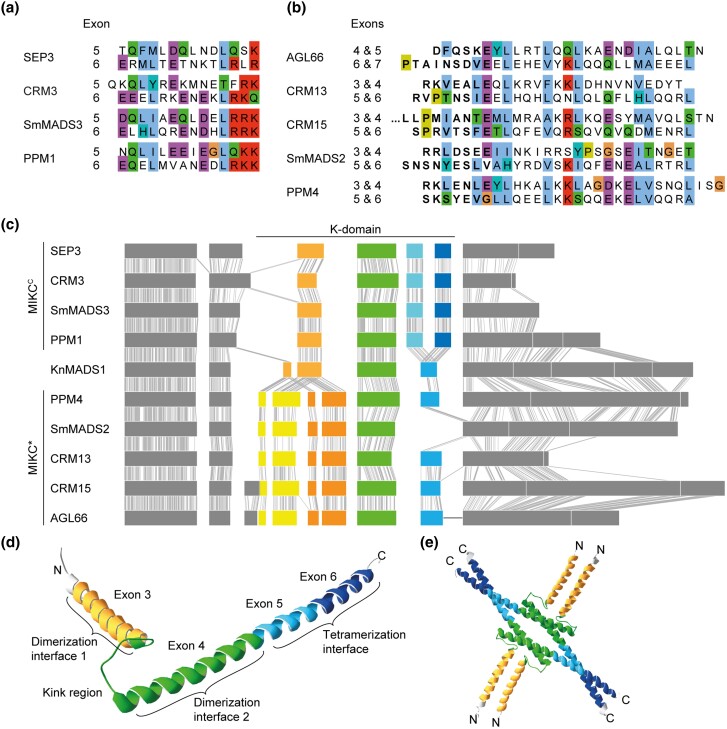
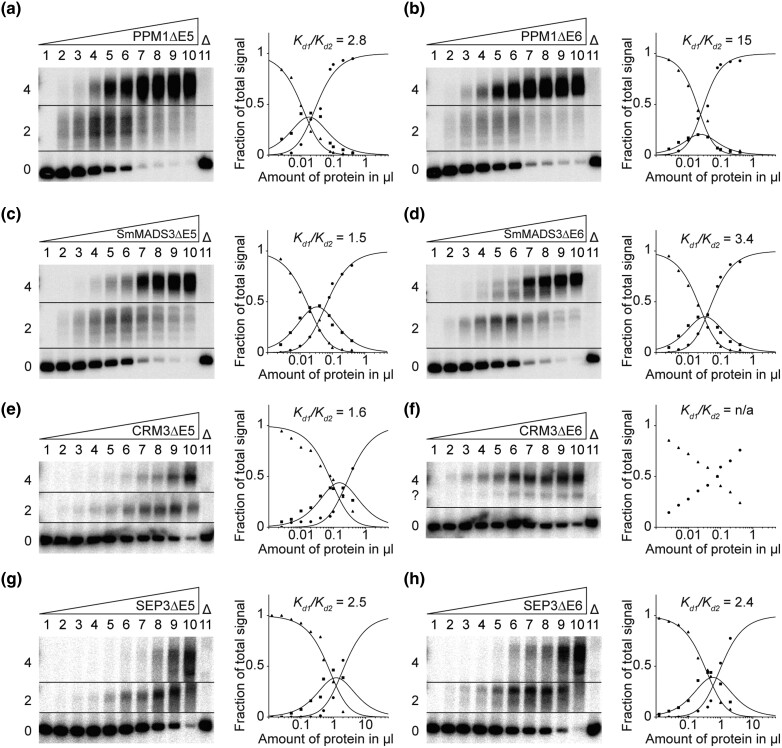
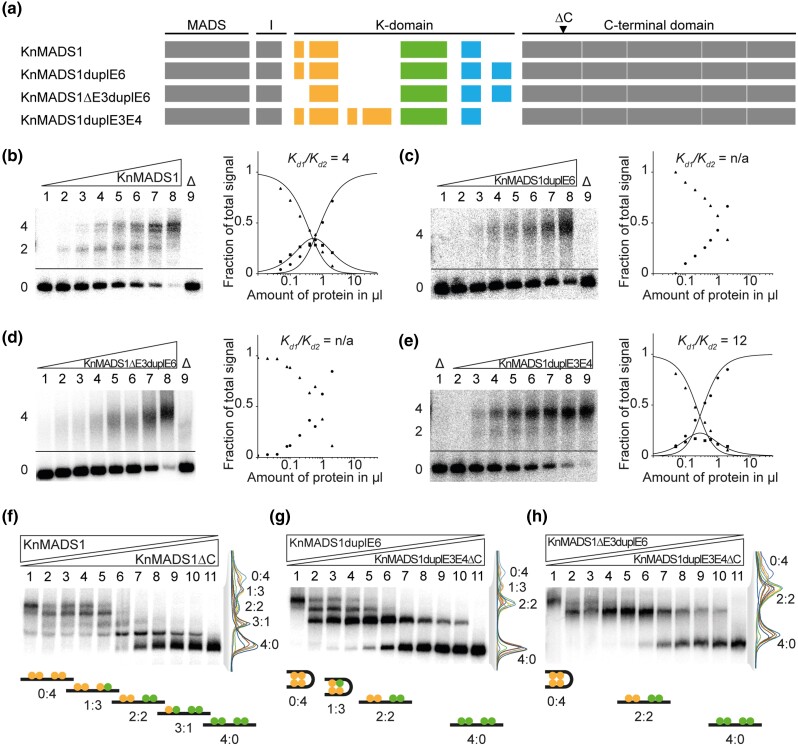
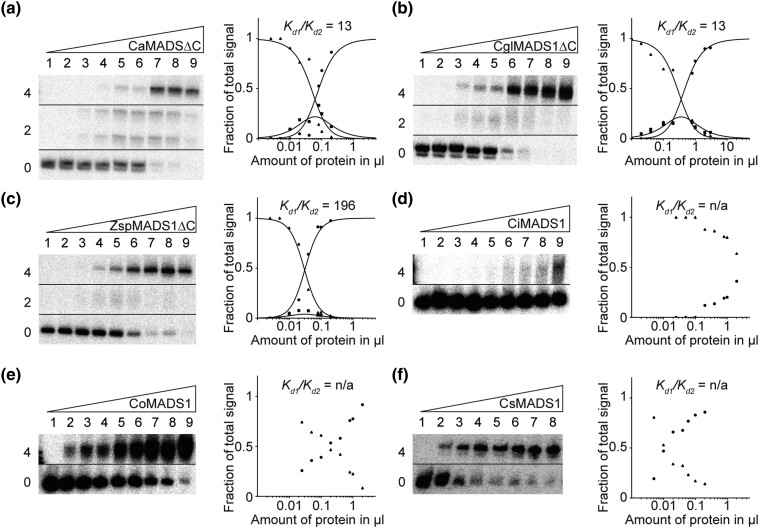
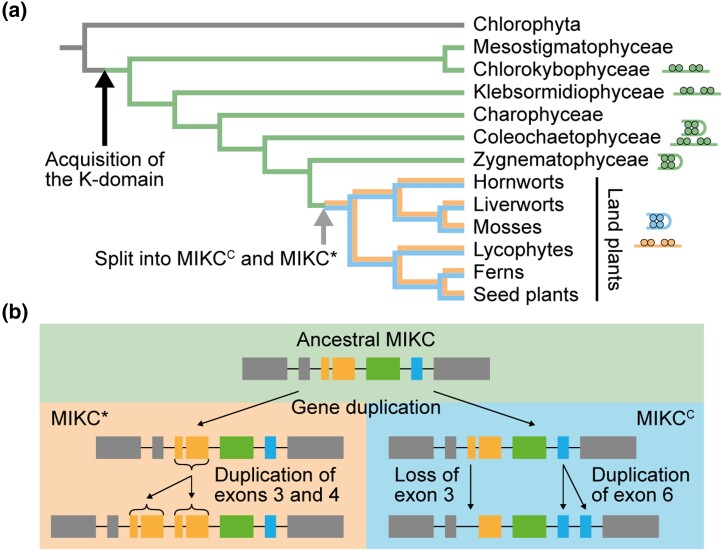
During development of flowering plants, some MIKC-type MADS-domain transcription factors (MTFs) exert their regulatory function as heterotetrameric complexes bound to two sites on the DNA of target genes. This way they constitute "floral quartets" or related "floral quartet-like complexes" (FQCs), involving a unique multimeric system of paralogous protein interactions. Tetramerization of MTFs is brought about mainly by interactions of keratin-like (K) domains. The K-domain associated with the more ancient DNA-binding MADS-domain during evolution in the stem group of extant streptophytes (charophyte green algae + land plants). However, whether this was sufficient for MTF tetramerization and FQC formation to occur, remains unknown. Here, we provide biophysical and bioinformatic data indicating that FQC formation likely originated in the stem group of land plants in a sublineage of MIKC-type genes termed MIKCC-type genes. In the stem group of this gene lineage, the duplication of the most downstream exon encoding the K-domain led to a C-terminal elongation of the second K-domain helix, thus, generating the tetramerization interface found in extant MIKCC-type proteins. In the stem group of the sister lineage of the MIKCC-type genes, termed MIKC*-type genes, the duplication of two other K-domain exons occurred, extending the K-domain at its N-terminal end. Our data indicate that this structural change prevents heterodimerization between MIKCC-type and MIKC*-type proteins. This way, two largely independent gene regulatory networks could be established, featuring MIKCC-type or MIKC*-type proteins, respectively, that control different aspects of plant development.
Keywords: MADS-box gene; MIKC-type MADS-domain transcription factor; cooperative DNA binding; floral quartet; keratin-like domain; protein–protein interaction.
© The Author(s) 2023. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Conflict of interest statement
Conflict of Interest statement. The authors declare no conflict of interest.
Figures
Similar articles
-
Cracking the Floral Quartet Code: How Do Multimers of MIKCC-Type MADS-Domain Transcription Factors Recognize Their Target Genes?Int J Mol Sci. 2023 May 4;24(9):8253. doi: 10.3390/ijms24098253. Int J Mol Sci. 2023. PMID: 37175955 Free PMC article. Review.
-
Two ancient classes of MIKC-type MADS-box genes are present in the moss Physcomitrella patens.Mol Biol Evol. 2002 Jun;19(6):801-14. doi: 10.1093/oxfordjournals.molbev.a004137. Mol Biol Evol. 2002. PMID: 12032236
-
How MIKC* MADS-box genes originated and evidence for their conserved function throughout the evolution of vascular plant gametophytes.Mol Biol Evol. 2012 Jan;29(1):293-302. doi: 10.1093/molbev/msr200. Epub 2011 Aug 3. Mol Biol Evol. 2012. PMID: 21813465
-
Novel members of the AGAMOUS LIKE 6 subfamily of MIKCC-type MADS-box genes in soybean.BMC Plant Biol. 2013 Jul 20;13:105. doi: 10.1186/1471-2229-13-105. BMC Plant Biol. 2013. PMID: 23870482 Free PMC article.
-
MIKC-type MADS-domain proteins: structural modularity, protein interactions and network evolution in land plants.Gene. 2005 Mar 14;347(2):183-98. doi: 10.1016/j.gene.2004.12.014. Epub 2005 Feb 22. Gene. 2005. PMID: 15777618 Review.
Cited by
-
Updated Phylogeny and Protein Structure Predictions Revise the Hypothesis on the Origin of MADS-box Transcription Factors in Land Plants.Mol Biol Evol. 2023 Sep 1;40(9):msad194. doi: 10.1093/molbev/msad194. Mol Biol Evol. 2023. PMID: 37652031 Free PMC article.
-
Cracking the Floral Quartet Code: How Do Multimers of MIKCC-Type MADS-Domain Transcription Factors Recognize Their Target Genes?Int J Mol Sci. 2023 May 4;24(9):8253. doi: 10.3390/ijms24098253. Int J Mol Sci. 2023. PMID: 37175955 Free PMC article. Review.
-
SEPALLATA-driven MADS transcription factor tetramerization is required for inner whorl floral organ development.Plant Cell. 2024 Sep 3;36(9):3435-3450. doi: 10.1093/plcell/koae151. Plant Cell. 2024. PMID: 38771250 Free PMC article.
-
Reflections on the ABC model of flower development.Plant Cell. 2024 May 1;36(5):1334-1357. doi: 10.1093/plcell/koae044. Plant Cell. 2024. PMID: 38345422 Free PMC article. Review.
-
Genome-wide identification and expression pattern analysis of MIKC-Type MADS-box genes in Chionanthus retusus, an androdioecy plant.BMC Genomics. 2024 Jul 2;25(1):662. doi: 10.1186/s12864-024-10569-8. BMC Genomics. 2024. PMID: 38956488 Free PMC article.
References
-
- Amoutzias GD, Robertson DL, Van de Peer Y, Oliver SG. 2008. Choose your partners: dimerization in eukaryotic transcription factors. Trends Biochem Sci. 33:220–229. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
