

PI(4,5)P2 regulates the gating of NaV1.4 channels
- PMID: 37043561
- PMCID: PMC10103707
- DOI: 10.1085/jgp.202213255
PI(4,5)P2 regulates the gating of NaV1.4 channels
Abstract
Voltage-gated sodium (NaV) channels are densely expressed in most excitable cells and activate in response to depolarization, causing a rapid influx of Na+ ions that initiates the action potential. The voltage-dependent activation of NaV channels is followed almost instantaneously by fast inactivation, setting the refractory period of excitable tissues. The gating cycle of NaV channels is subject to tight regulation, with perturbations leading to a range of pathophysiological states. The gating properties of most ion channels are regulated by the membrane phospholipid, phosphatidylinositol (4,5) bisphosphate (PI(4,5)P2). However, it is not known whether PI(4,5)P2 modulates the activity of NaV channels. Here, we utilize optogenetics to activate specific, membrane-associated phosphoinositide (PI)-phosphatases that dephosphorylate PI(4,5)P2 while simultaneously recording NaV1.4 channel currents. We show that dephosphorylating PI(4,5)P2 left-shifts the voltage-dependent gating of NaV1.4 to more hyperpolarized membrane potentials, augments the late current that persists after fast inactivation, and speeds the rate at which channels recover from fast inactivation. These effects are opposed by exogenous diC8PI(4,5)P2. We provide evidence that PI(4,5)P2 is a negative regulator that tunes the gating behavior of NaV1.4 channels.
© 2023 Gada et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures


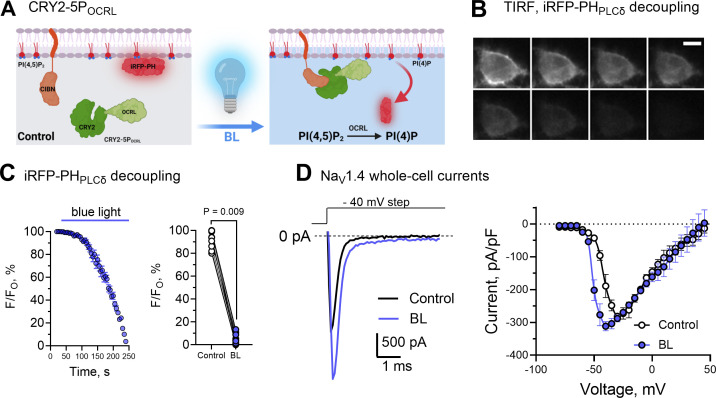



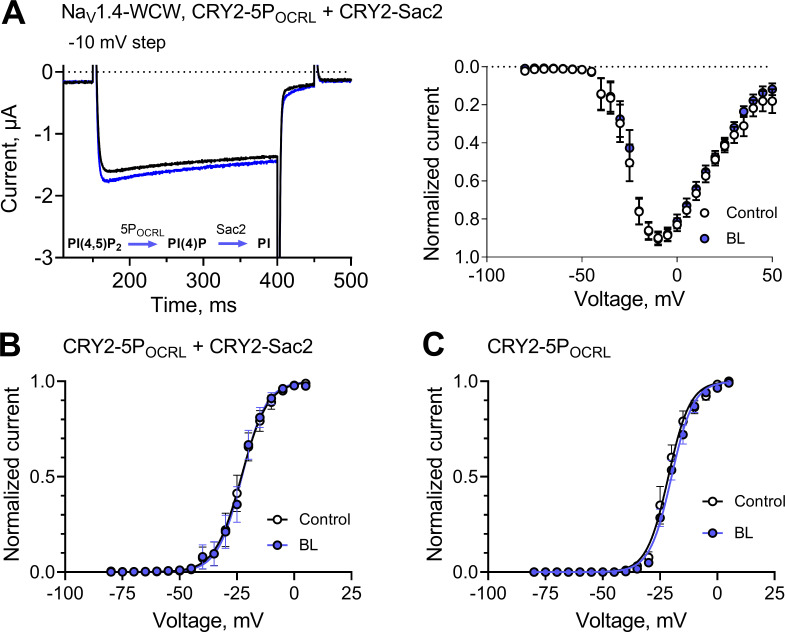
Comment in
-
Phosphoinositide regulation of voltage-gated sodium channels.J Gen Physiol. 2023 Jun 5;155(6):e202313402. doi: 10.1085/jgp.202313402. Epub 2023 May 11. J Gen Physiol. 2023. PMID: 37166350 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
