

The ins and outs of innate and adaptive type 2 immunity
- PMID: 37044061
- PMCID: PMC10120575
- DOI: 10.1016/j.immuni.2023.03.014
The ins and outs of innate and adaptive type 2 immunity
Abstract
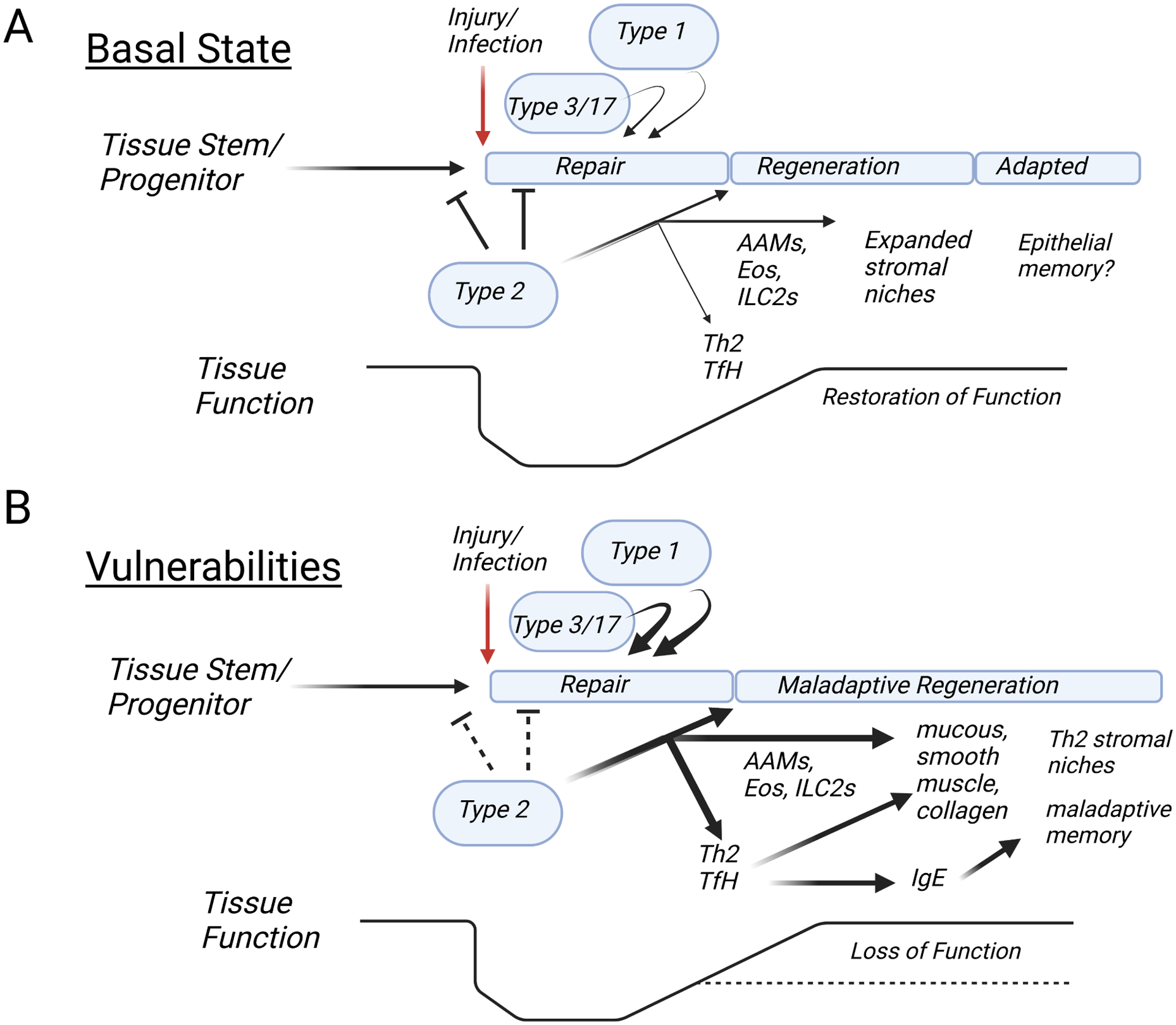
Type 2 immunity is orchestrated by a canonical group of cytokines primarily produced by innate lymphoid cells, group 2, and their adaptive counterparts, CD4+ helper type 2 cells, and elaborated by myeloid cells and antibodies that accumulate in response. Here, we review the cytokine and cellular circuits that mediate type 2 immunity. Building from insights in cytokine evolution, we propose that innate type 2 immunity evolved to monitor the status of microbe-rich epithelial barriers (outside) and sterile parenchymal borders (inside) to meet the functional demands of local tissue, and, when necessary, to relay information to the adaptive immune system to reinforce demarcating borders to sustain these efforts. Allergic pathology likely results from deviations in local sustaining units caused by alterations imposed by environmental effects during postnatal developmental windows and exacerbated by mutations that increase vulnerabilities. This framework positions T2 immunity as central to sustaining tissue repair and regeneration and provides a context toward understanding allergic disease.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests R.M.L. is a member of the Scientific Advisory Board for Genentech and a participant on the Editorial Advisory Board at Immunity.
Figures
References
-
- Hammad H, Lambrecht BN. 2021. The basic immunology of asthma. Cell 184:1469–1485. - PubMed
-
- Sharma M, Leung D, Momenilandi M, Jones LCW, Pacillo L, James AE, Murrell JR, Delafontaine S, Maimaris J, Vaseghi-Shanjani M, Del Bel KL, Lu HY, Chua GT, Di Cesare S, Fornes O, Liu Z, De Matteo G, Fu MP, Amodio D, et al. 2023. Human germline heterozygous gain-of-function STAT6 variants cause severe allergic disease. J Exp Med 220:20221437. - PMC - PubMed
