

Ageing-associated changes in transcriptional elongation influence longevity
- PMID: 37046086
- PMCID: PMC10132977
- DOI: 10.1038/s41586-023-05922-y
Ageing-associated changes in transcriptional elongation influence longevity
Abstract
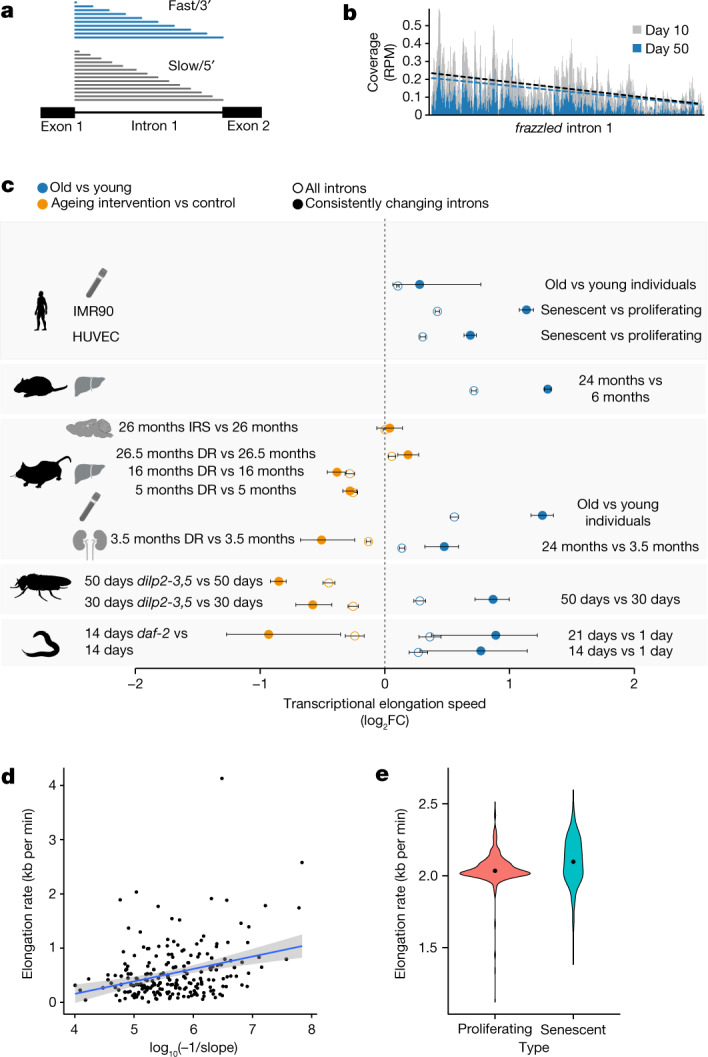
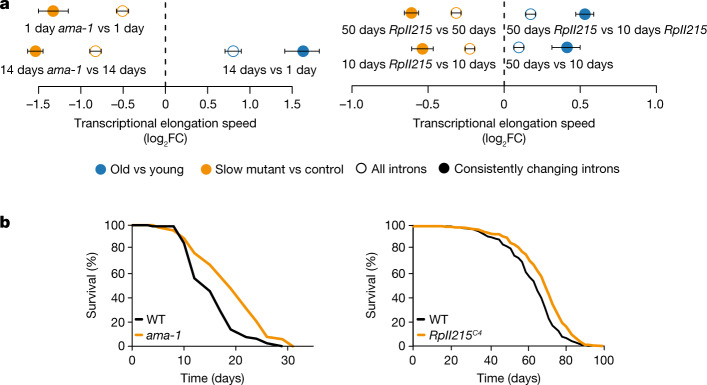
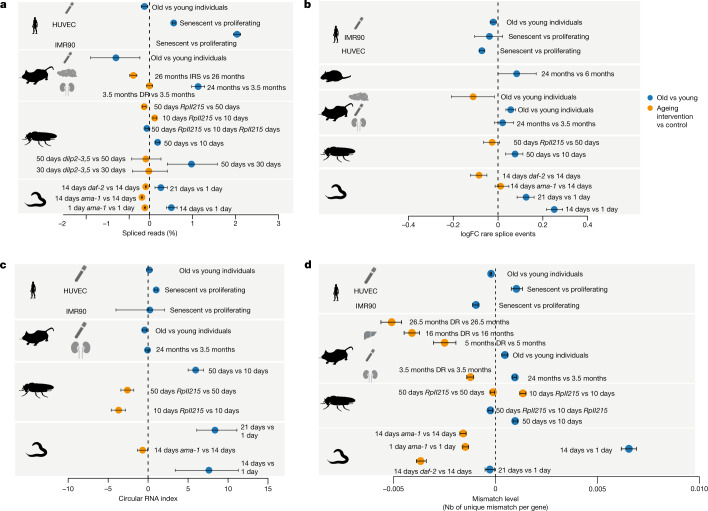
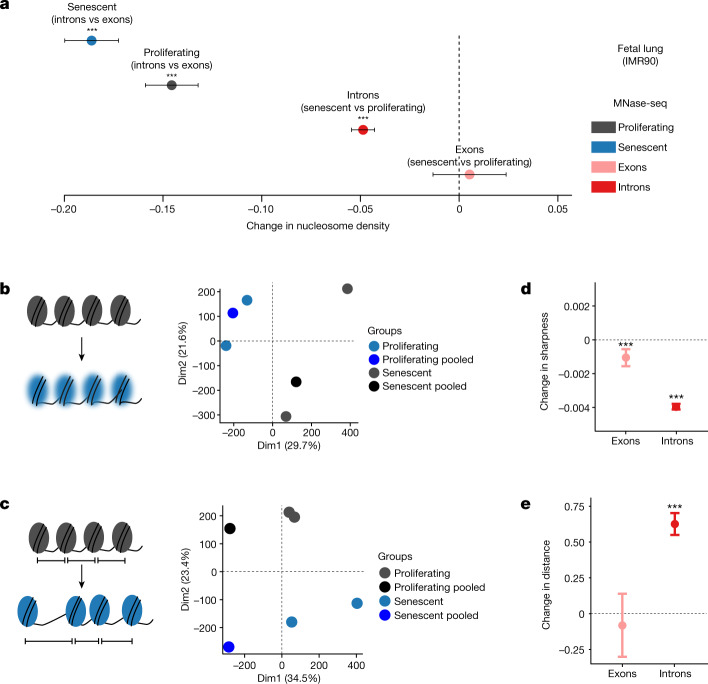
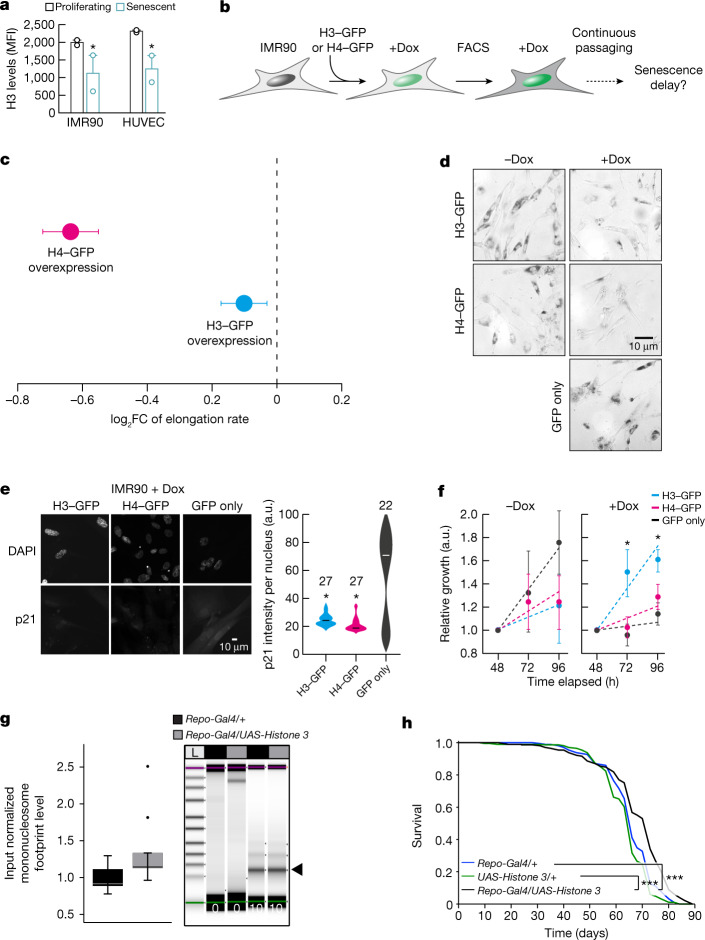
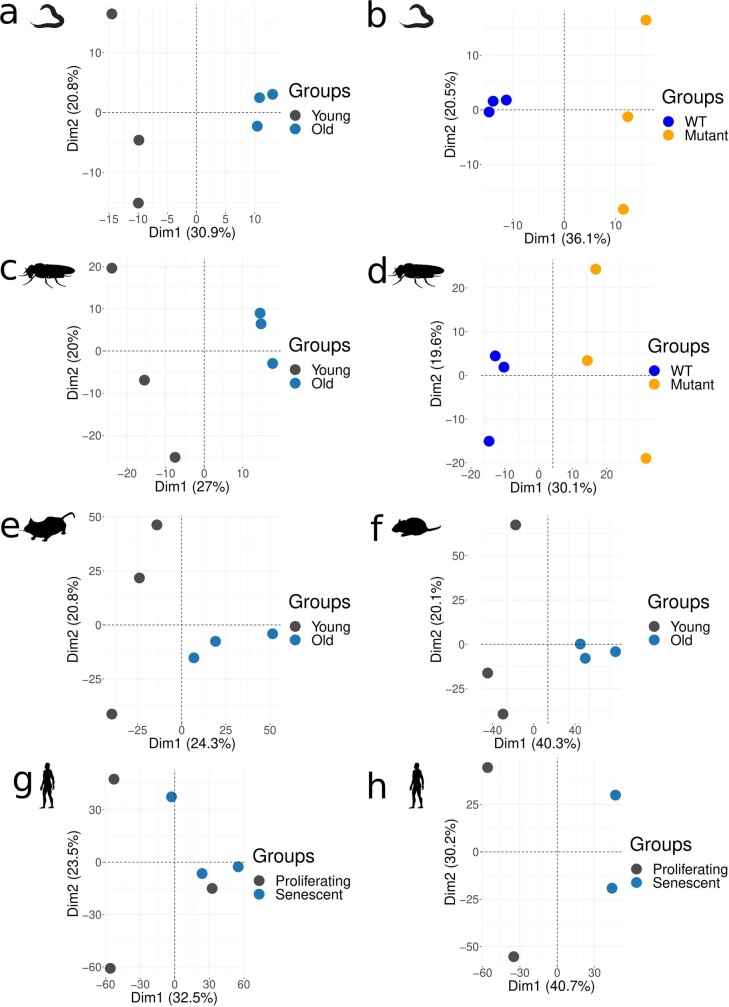
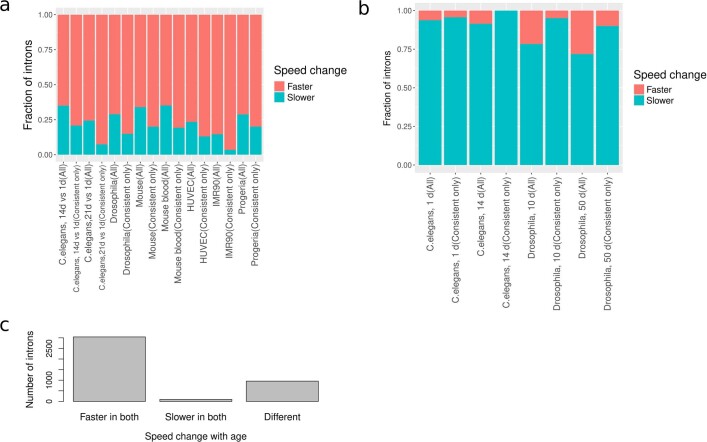
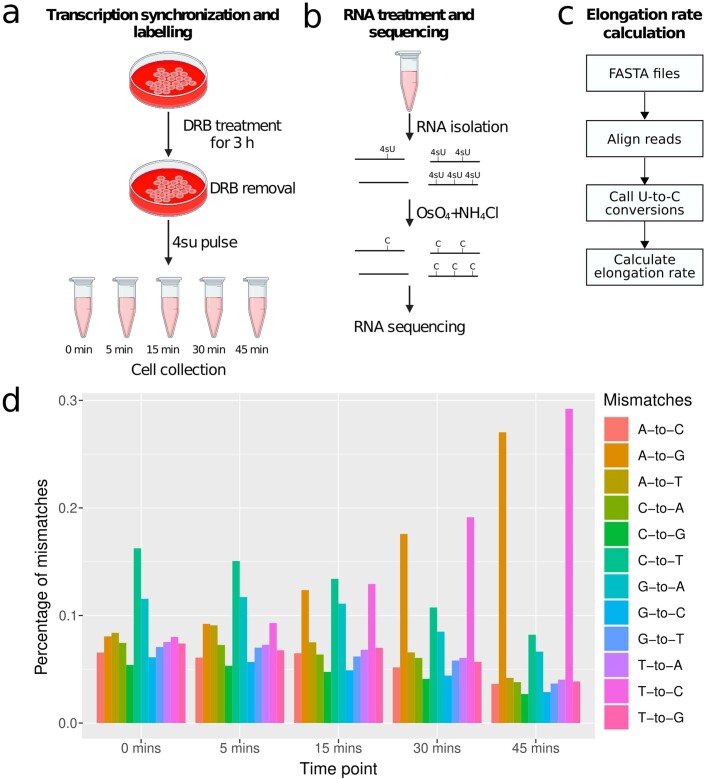
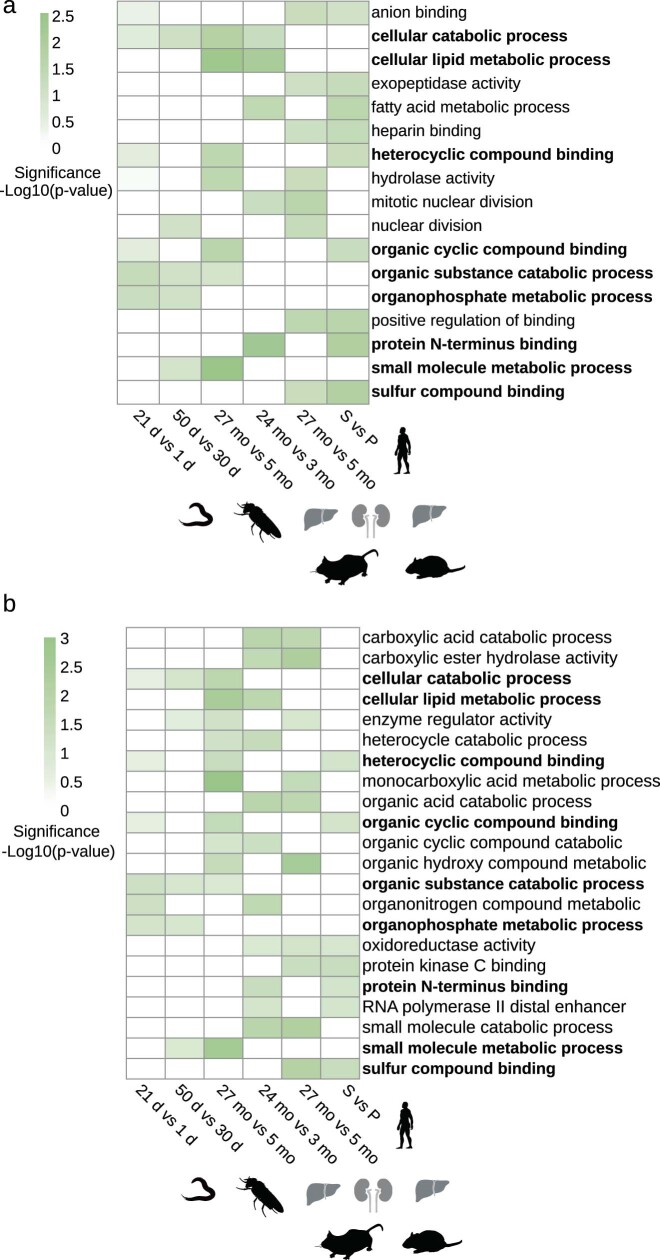
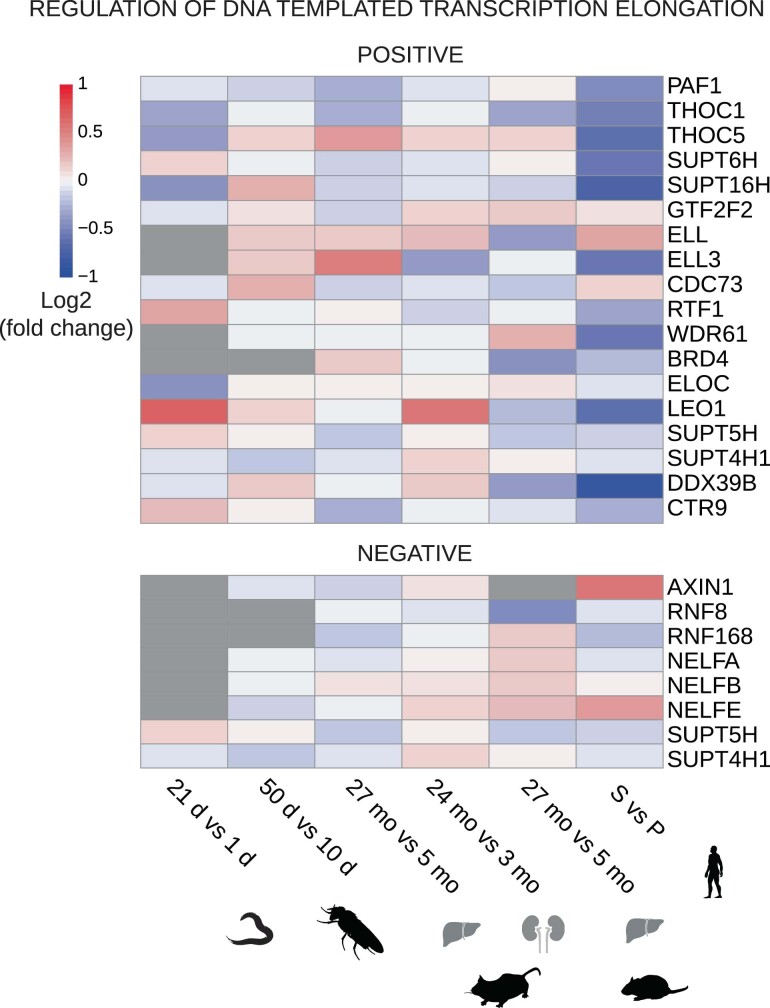
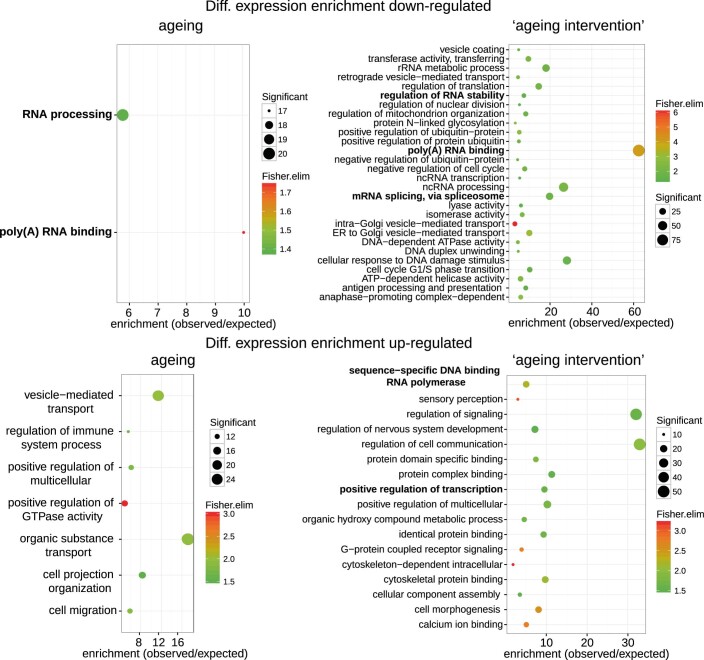
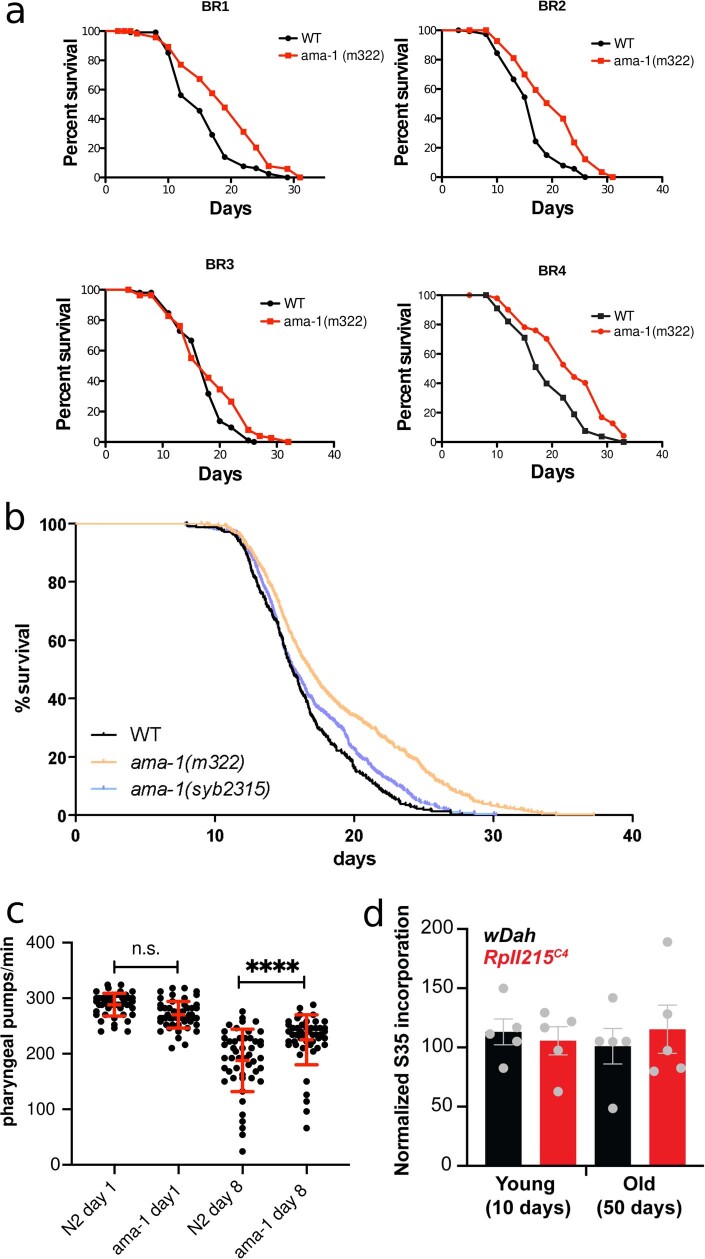
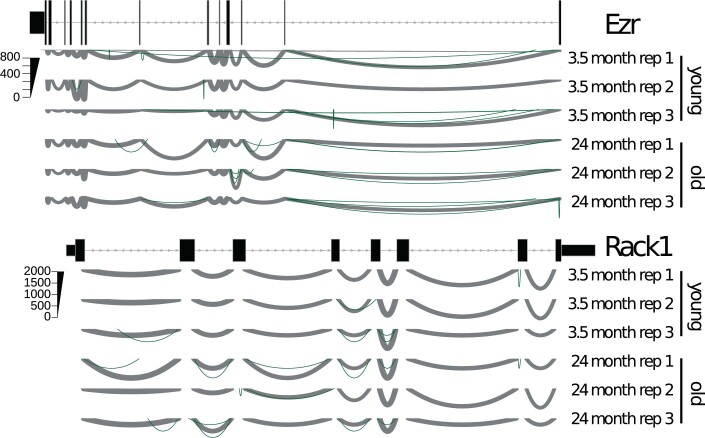
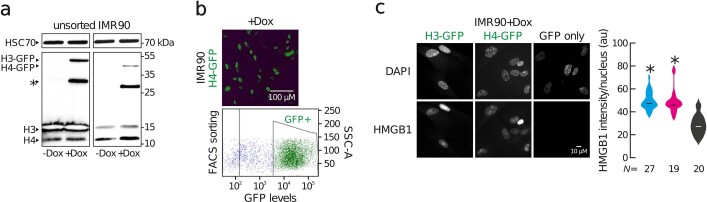
Physiological homeostasis becomes compromised during ageing, as a result of impairment of cellular processes, including transcription and RNA splicing1-4. However, the molecular mechanisms leading to the loss of transcriptional fidelity are so far elusive, as are ways of preventing it. Here we profiled and analysed genome-wide, ageing-related changes in transcriptional processes across different organisms: nematodes, fruitflies, mice, rats and humans. The average transcriptional elongation speed (RNA polymerase II speed) increased with age in all five species. Along with these changes in elongation speed, we observed changes in splicing, including a reduction of unspliced transcripts and the formation of more circular RNAs. Two lifespan-extending interventions, dietary restriction and lowered insulin-IGF signalling, both reversed most of these ageing-related changes. Genetic variants in RNA polymerase II that reduced its speed in worms5 and flies6 increased their lifespan. Similarly, reducing the speed of RNA polymerase II by overexpressing histone components, to counter age-associated changes in nucleosome positioning, also extended lifespan in flies and the division potential of human cells. Our findings uncover fundamental molecular mechanisms underlying animal ageing and lifespan-extending interventions, and point to possible preventive measures.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Comment in
-
Aging RNA Polymerase II may speed recklessly.Nat Struct Mol Biol. 2023 May;30(5):567. doi: 10.1038/s41594-023-01005-8. Nat Struct Mol Biol. 2023. PMID: 37198270 No abstract available.
-
Accelerated transcriptional elongation during aging impairs longevity.Cell Res. 2023 Nov;33(11):817-818. doi: 10.1038/s41422-023-00829-9. Cell Res. 2023. PMID: 37253838 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
