

Dynamic synchronization between hippocampal representations and stepping
- PMID: 37046088
- PMCID: PMC10156593
- DOI: 10.1038/s41586-023-05928-6
Dynamic synchronization between hippocampal representations and stepping
Abstract
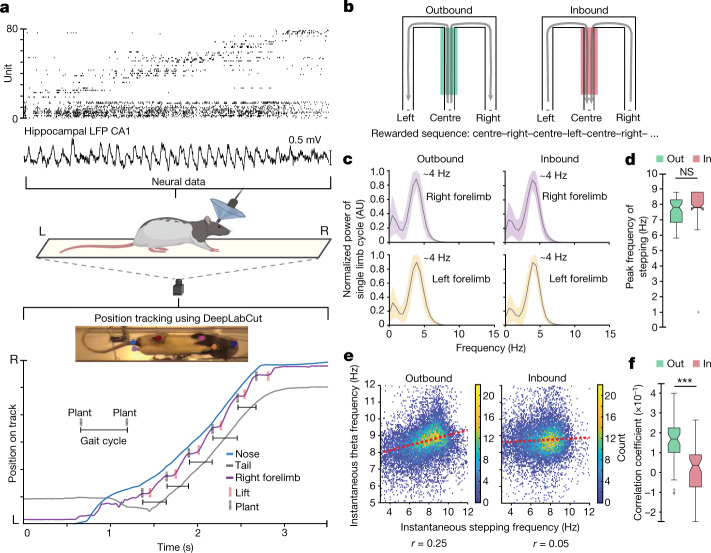
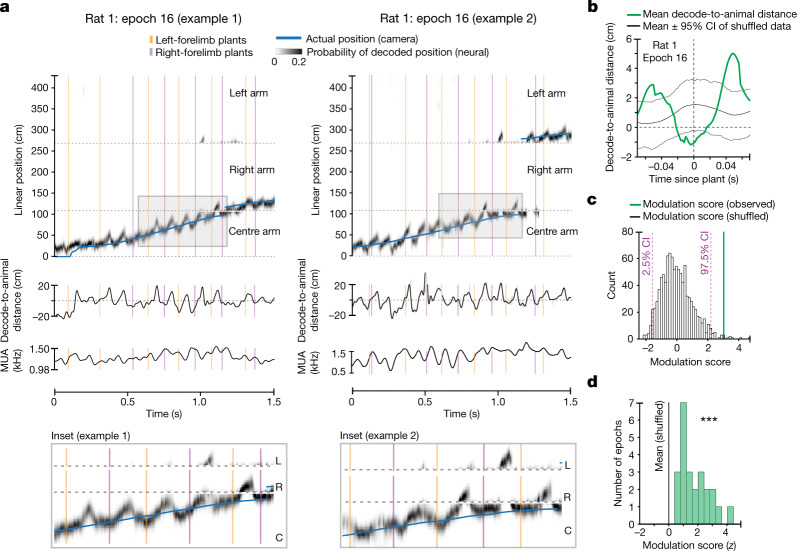
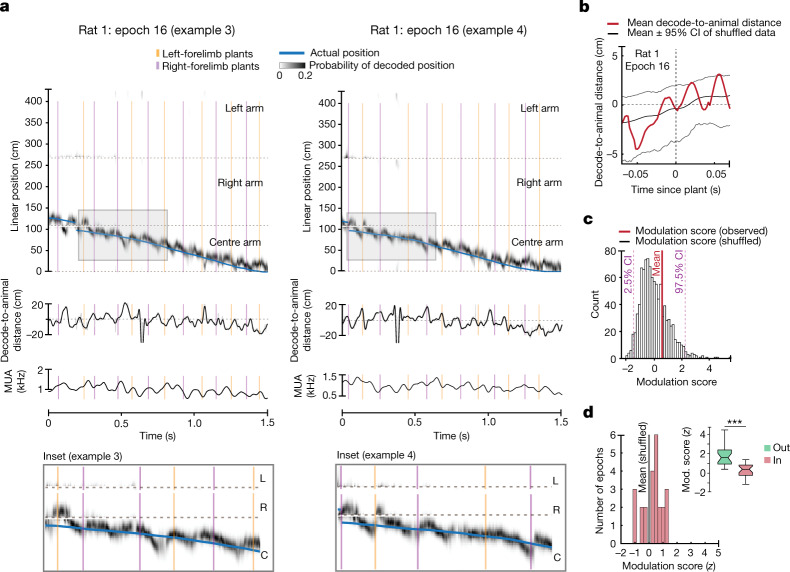
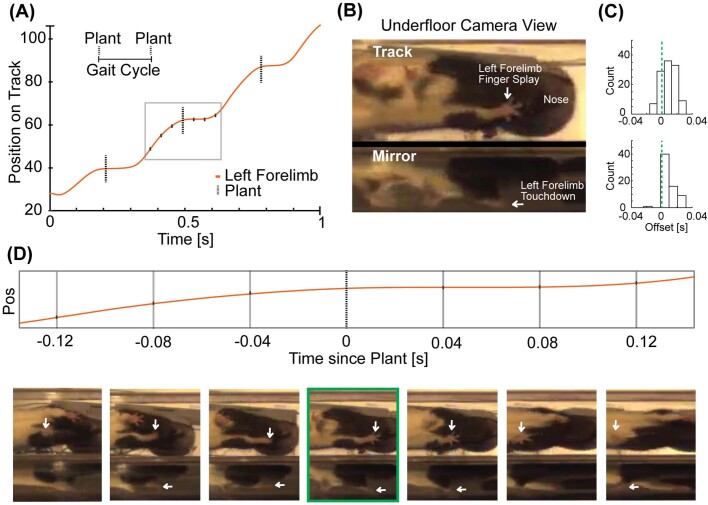
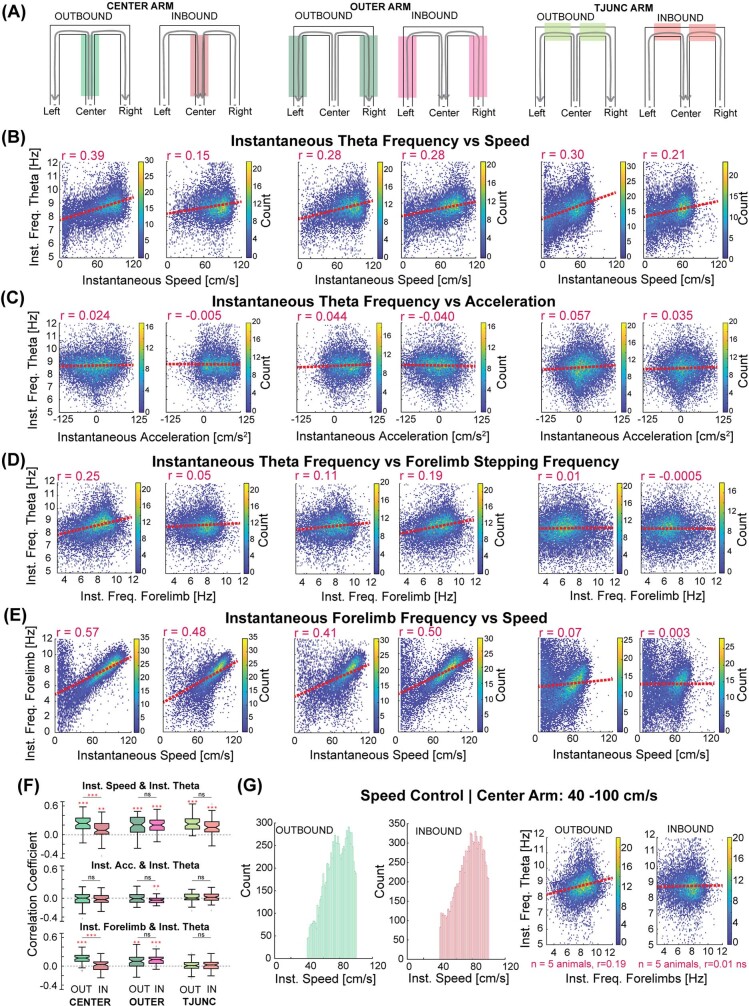
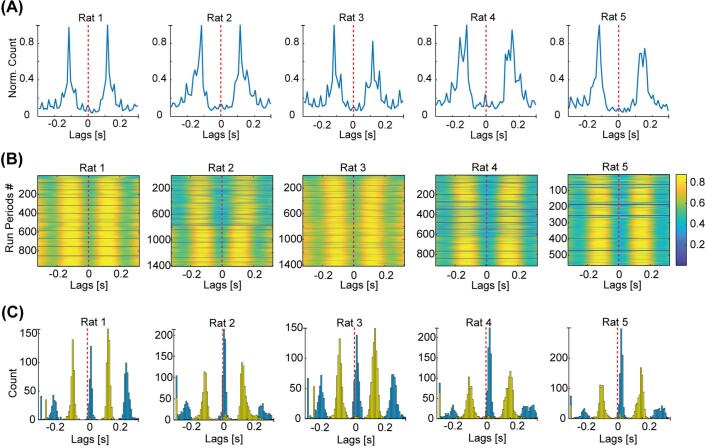
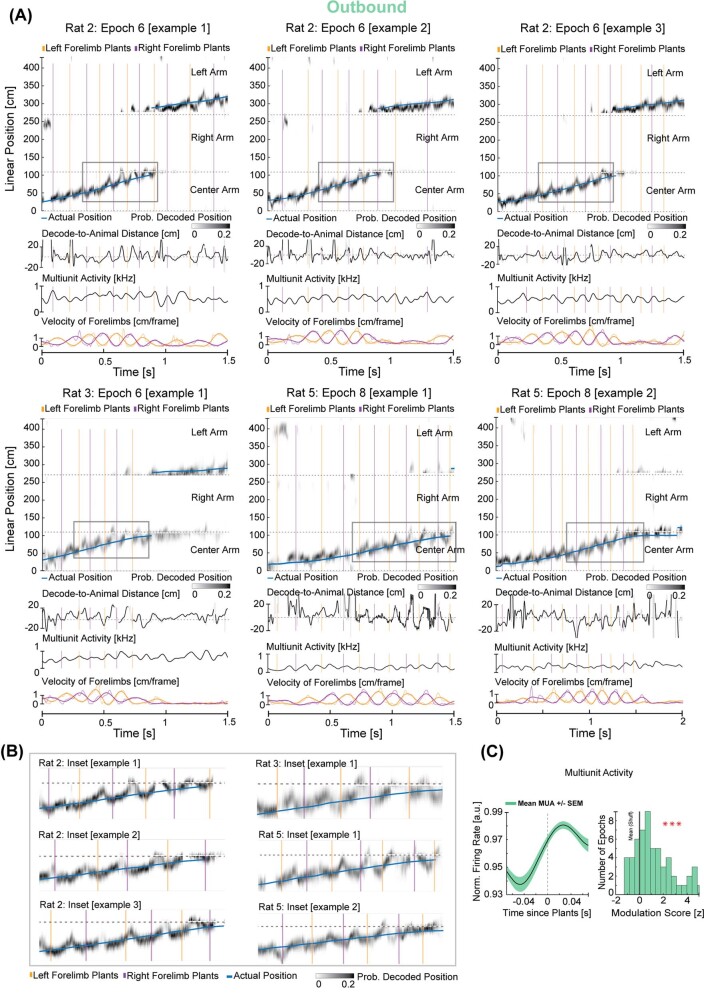
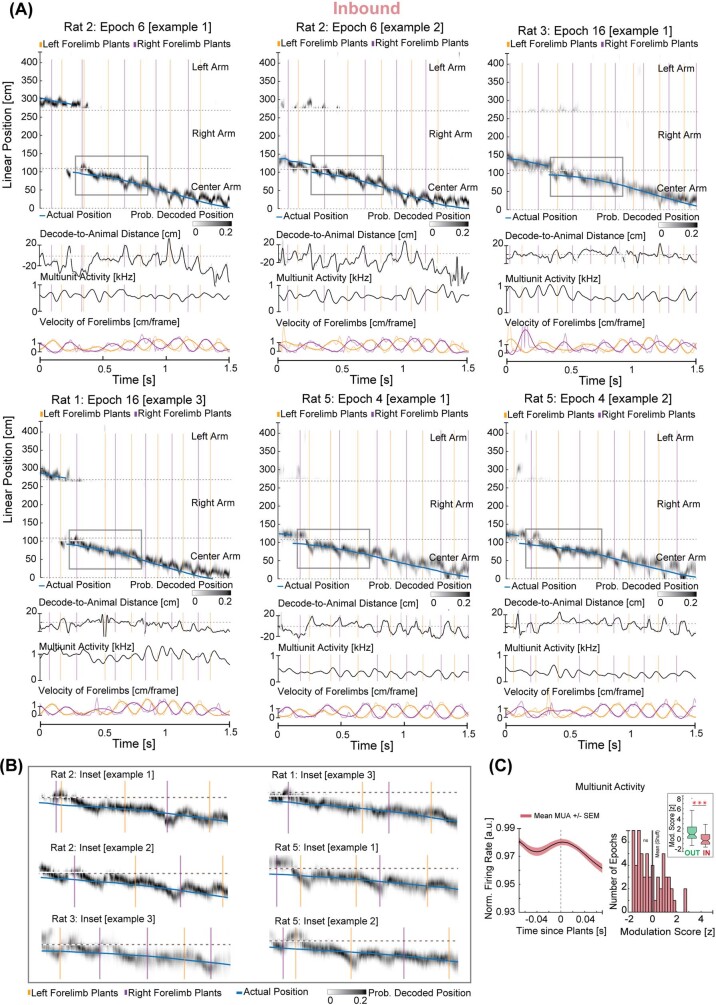
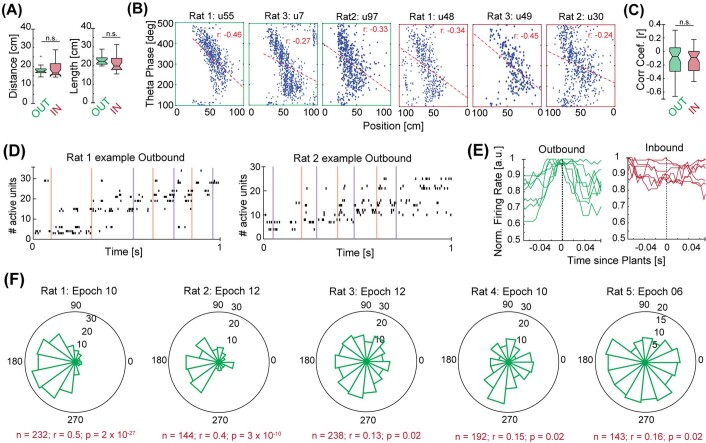
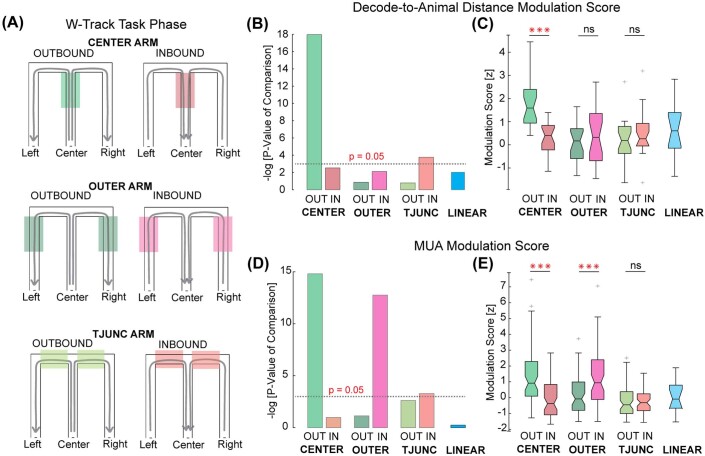
The hippocampus is a mammalian brain structure that expresses spatial representations1 and is crucial for navigation2,3. Navigation, in turn, intricately depends on locomotion; however, current accounts suggest a dissociation between hippocampal spatial representations and the details of locomotor processes. Specifically, the hippocampus is thought to represent mainly higher-order cognitive and locomotor variables such as position, speed and direction of movement4-7, whereas the limb movements that propel the animal can be computed and represented primarily in subcortical circuits, including the spinal cord, brainstem and cerebellum8-11. Whether hippocampal representations are actually decoupled from the detailed structure of locomotor processes remains unknown. To address this question, here we simultaneously monitored hippocampal spatial representations and ongoing limb movements underlying locomotion at fast timescales. We found that the forelimb stepping cycle in freely behaving rats is rhythmic and peaks at around 8 Hz during movement, matching the approximately 8 Hz modulation of hippocampal activity and spatial representations during locomotion12. We also discovered precisely timed coordination between the time at which the forelimbs touch the ground ('plant' times of the stepping cycle) and the hippocampal representation of space. Notably, plant times coincide with hippocampal representations that are closest to the actual position of the nose of the rat, whereas between these plant times, the hippocampal representation progresses towards possible future locations. This synchronization was specifically detectable when rats approached spatial decisions. Together, our results reveal a profound and dynamic coordination on a timescale of tens of milliseconds between central cognitive representations and peripheral motor processes. This coordination engages and disengages rapidly in association with cognitive demands and is well suited to support rapid information exchange between cognitive and sensory-motor circuits.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Descending pathways eliciting forelimb stepping in the lateral funiculus: experimental studies with stimulation and lesion of the cervical cord in decerebrate cats.Brain Res. 1986 Jul 30;379(1):125-36. doi: 10.1016/0006-8993(86)90264-7. Brain Res. 1986. PMID: 3742207
-
The ventral hippocampus is involved in multi-goal obstacle-rich spatial navigation.Hippocampus. 2018 Dec;28(12):853-866. doi: 10.1002/hipo.22993. Epub 2018 Nov 5. Hippocampus. 2018. PMID: 30067283
-
Hippocampal gamma rhythms during Y-maze navigation in the juvenile rat.Hippocampus. 2020 May;30(5):505-525. doi: 10.1002/hipo.23168. Epub 2019 Oct 18. Hippocampus. 2020. PMID: 31626396 Free PMC article.
-
Navigating cognition: Spatial codes for human thinking.Science. 2018 Nov 9;362(6415):eaat6766. doi: 10.1126/science.aat6766. Science. 2018. PMID: 30409861 Review.
-
Spatial cognition in bats and rats: from sensory acquisition to multiscale maps and navigation.Nat Rev Neurosci. 2015 Feb;16(2):94-108. doi: 10.1038/nrn3888. Nat Rev Neurosci. 2015. PMID: 25601780 Review.
Cited by
-
Electrical stimulation: a potential alternative to positively impact cerebral health?Front Physiol. 2024 Sep 20;15:1464326. doi: 10.3389/fphys.2024.1464326. eCollection 2024. Front Physiol. 2024. PMID: 39371600 Free PMC article. Review.
-
Tactile Mechanisms and Afferents Underlying the Rat Pup Transport Response.bioRxiv [Preprint]. 2024 Aug 23:2024.08.23.609194. doi: 10.1101/2024.08.23.609194. bioRxiv. 2024. Update in: Curr Biol. 2024 Dec 2;34(23):5595-5601.e2. doi: 10.1016/j.cub.2024.10.016. PMID: 39229029 Free PMC article. Updated. Preprint.
-
Fantastic beasts and how to study them: rethinking experimental animal behavior.J Exp Biol. 2024 Feb 15;227(4):jeb247003. doi: 10.1242/jeb.247003. Epub 2024 Feb 19. J Exp Biol. 2024. PMID: 38372042 Free PMC article.
-
Behavioral timescale synaptic plasticity in the hippocampus creates non-spatial representations during learning and is modulated by entorhinal inputs.bioRxiv [Preprint]. 2025 Apr 1:2024.08.27.609983. doi: 10.1101/2024.08.27.609983. bioRxiv. 2025. PMID: 39253411 Free PMC article. Preprint.
-
A Hippocampal-Parietal Network for Reference Frame Coordination.J Neurosci. 2025 Apr 23;45(17):e1782242025. doi: 10.1523/JNEUROSCI.1782-24.2025. J Neurosci. 2025. PMID: 39909564
