

Recent Theoretical Insights into the Oxidative Degradation of Biopolymers and Plastics by Metalloenzymes
- PMID: 37047341
- PMCID: PMC10094197
- DOI: 10.3390/ijms24076368
Recent Theoretical Insights into the Oxidative Degradation of Biopolymers and Plastics by Metalloenzymes
Abstract
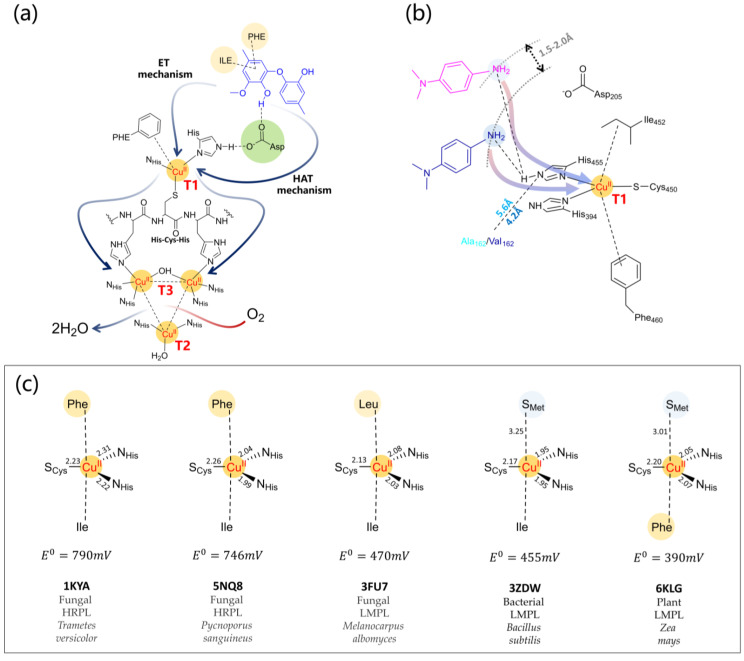
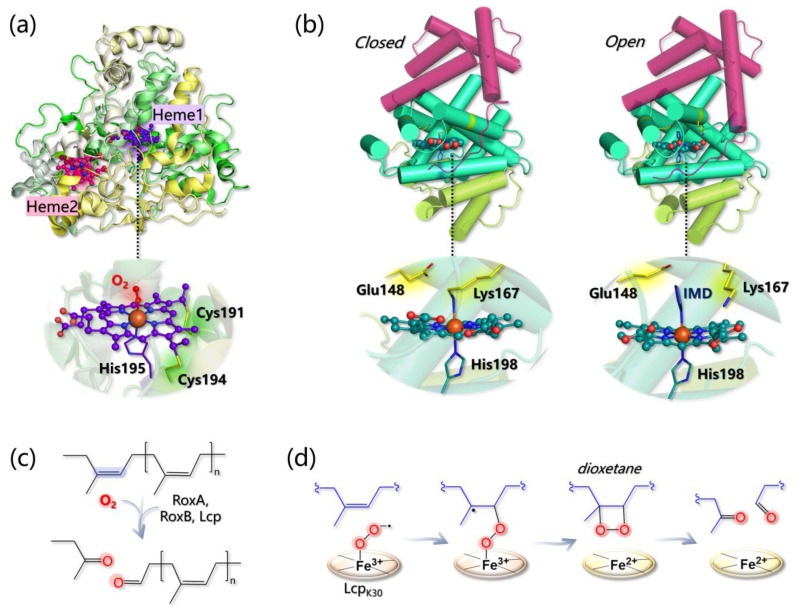
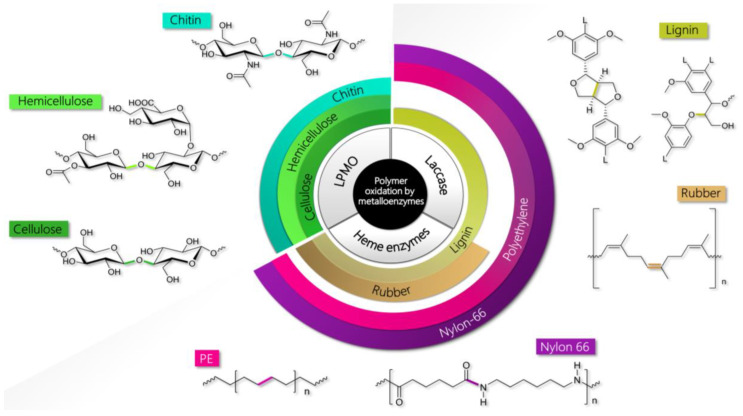
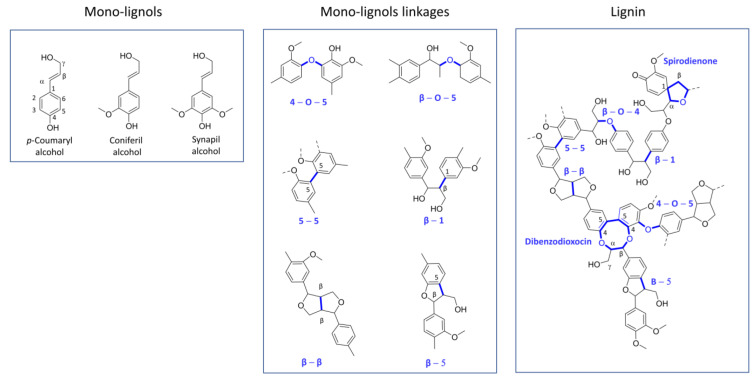
Molecular modeling techniques have become indispensable in many fields of molecular sciences in which the details related to mechanisms and reactivity need to be studied at an atomistic level. This review article provides a collection of computational modeling works on a topic of enormous interest and urgent relevance: the properties of metalloenzymes involved in the degradation and valorization of natural biopolymers and synthetic plastics on the basis of both circular biofuel production and bioremediation strategies. In particular, we will focus on lytic polysaccharide monooxygenase, laccases, and various heme peroxidases involved in the processing of polysaccharides, lignins, rubbers, and some synthetic polymers. Special attention will be dedicated to the interaction between these enzymes and their substrate studied at different levels of theory, starting from classical molecular docking and molecular dynamics techniques up to techniques based on quantum chemistry.
Keywords: biofuels; biopolymers; bioremediation; laccases; lytic polysaccharide monooxygenase; molecular modeling; oxidative metalloenzymes; peroxidases; plastic; rubber oxygenases.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
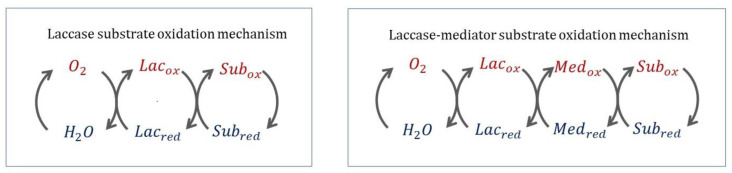
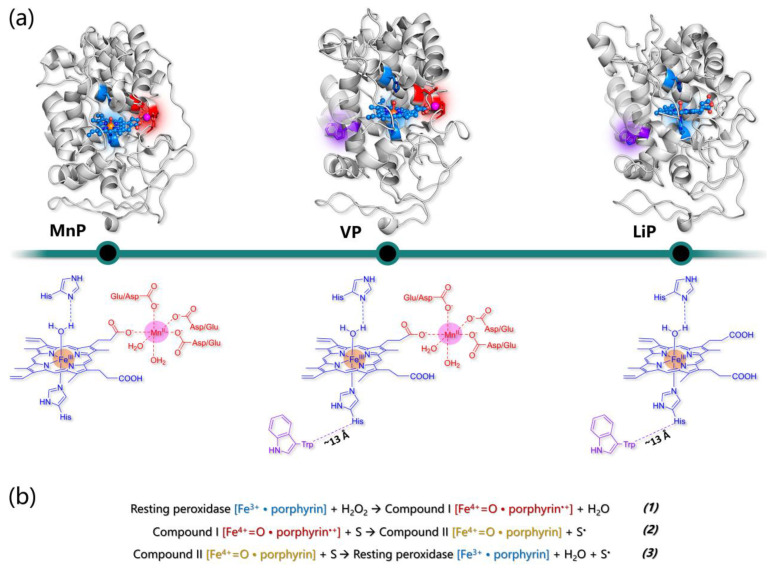
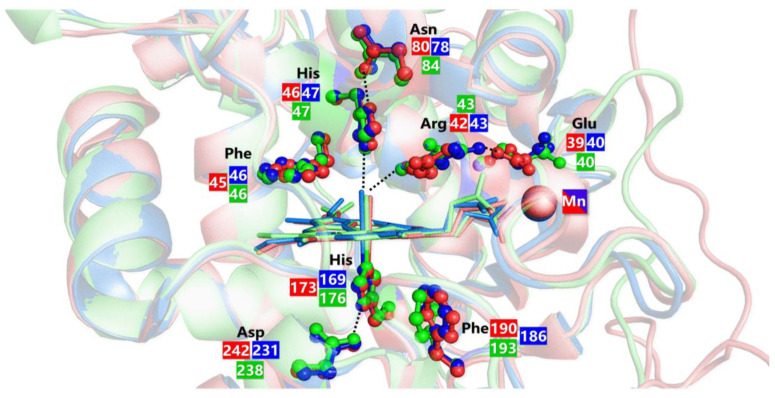
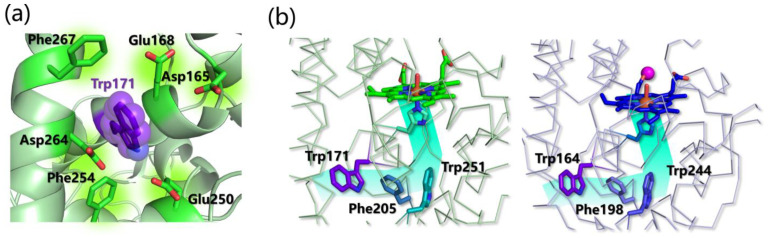
Similar articles
-
Insights into lignin degradation and its potential industrial applications.Adv Appl Microbiol. 2013;82:1-28. doi: 10.1016/B978-0-12-407679-2.00001-6. Adv Appl Microbiol. 2013. PMID: 23415151
-
Oxidoreductases on their way to industrial biotransformations.Biotechnol Adv. 2017 Nov 1;35(6):815-831. doi: 10.1016/j.biotechadv.2017.06.003. Epub 2017 Jun 15. Biotechnol Adv. 2017. PMID: 28624475 Review.
-
In situ H2 O2 Generation by Choline Oxidase and Its Application in Amino Polysaccharide Degradation by Coupling to Lytic Polysaccharide Monooxygenase.Chembiochem. 2023 Jul 17;24(14):e202300363. doi: 10.1002/cbic.202300363. Epub 2023 Jun 21. Chembiochem. 2023. PMID: 37191321
-
A Lytic Polysaccharide Monooxygenase from a White-Rot Fungus Drives the Degradation of Lignin by a Versatile Peroxidase.Appl Environ Microbiol. 2019 Apr 18;85(9):e02803-18. doi: 10.1128/AEM.02803-18. Print 2019 May 1. Appl Environ Microbiol. 2019. PMID: 30824433 Free PMC article.
-
Thermophilic enzyme systems for efficient conversion of lignocellulose to valuable products: Structural insights and future perspectives for esterases and oxidative catalysts.Bioresour Technol. 2019 May;279:362-372. doi: 10.1016/j.biortech.2019.01.062. Epub 2019 Jan 16. Bioresour Technol. 2019. PMID: 30685134 Review.
Cited by
-
In Vitro and Computational Response of Differential Catalysis by Phlebia brevispora BAFC 633 Laccase in Interaction with 2,4-D and Chlorpyrifos.Int J Mol Sci. 2024 Nov 22;25(23):12527. doi: 10.3390/ijms252312527. Int J Mol Sci. 2024. PMID: 39684240 Free PMC article.
-
Natural, Designed and Engineered Metalloenzymes: Structure, Catalytic Mechanisms and Applications.Int J Mol Sci. 2023 Sep 19;24(18):14255. doi: 10.3390/ijms241814255. Int J Mol Sci. 2023. PMID: 37762558 Free PMC article.
-
Responses of natural plastisphere community and zooplankton to microplastic pollution: a review on novel remediation strategies.Arch Microbiol. 2025 May 7;207(6):136. doi: 10.1007/s00203-025-04334-y. Arch Microbiol. 2025. PMID: 40332619 Review.
-
A novel laccase from Trametes polyzona with high performance in the decolorization of textile dyes.AMB Express. 2024 Mar 20;14(1):32. doi: 10.1186/s13568-024-01687-3. AMB Express. 2024. PMID: 38506984 Free PMC article.
-
Isolation and Characterization of Laccase from Trichoderma asperellum Tasjk65.Biology (Basel). 2025 Jun 13;14(6):691. doi: 10.3390/biology14060691. Biology (Basel). 2025. PMID: 40563942 Free PMC article.
References
-
- Osman A.I., Mehta N., Elgarahy A.M., Al-Hinai A., Al-Muhtaseb A.H., Rooney D.W. Conversion of Biomass to Biofuels and Life Cycle Assessment: A Review. Environ. Chem. Lett. 2021;19:4075–4118. doi: 10.1007/s10311-021-01273-0. - DOI
-
- Abdallah Q.A., Al Abdallah Q., Tracy Nixon B., Fortwendel J.R. The Enzymatic Conversion of Major Algal and Cyanobacterial Carbohydrates to Bioethanol. Front. Energy Res. 2016;4:36. doi: 10.3389/fenrg.2016.00036. - DOI
-
- Sweeney M.D., Xu F. Biomass Converting Enzymes as Industrial Biocatalysts for Fuels and Chemicals: Recent Developments. Catalysts. 2012;2:244–263. doi: 10.3390/catal2020244. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
