

TLR9 Signaling Is Required for the Porphyromonas gingivalis-Induced Activation of IL-10-Expressing B Cells
- PMID: 37047666
- PMCID: PMC10094902
- DOI: 10.3390/ijms24076693
TLR9 Signaling Is Required for the Porphyromonas gingivalis-Induced Activation of IL-10-Expressing B Cells
Abstract
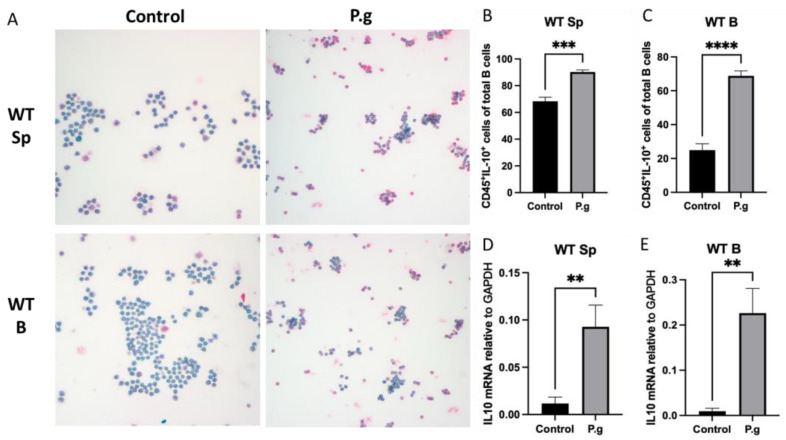
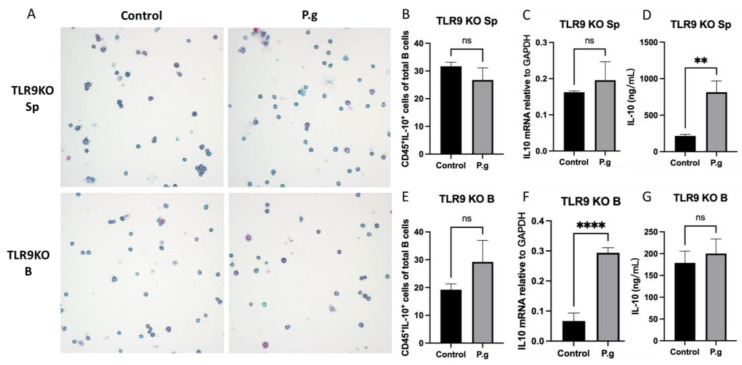
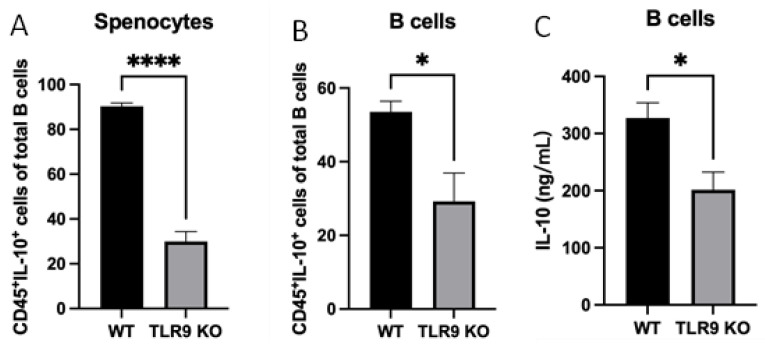
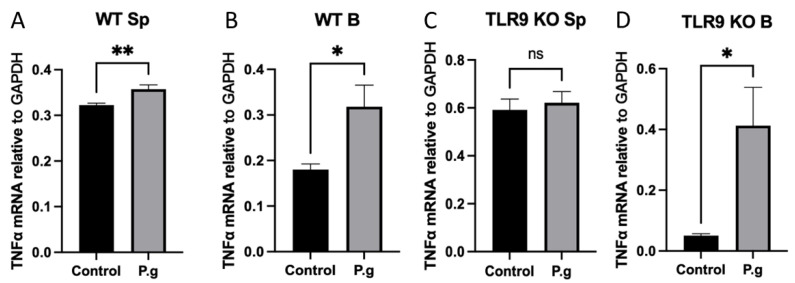
Immune cell pattern-recognition receptors such as Toll-like receptors (TLRs) play important roles in the regulation of host responses to periodontal pathogens. Our previous studies have demonstrated that immune regulatory B cells were activated by TLRs and alleviated periodontitis inflammation and bone loss. The purpose of this study is to determine the role of TLR9 signaling in the activation and IL-10 production of the primed-immune B cells in vitro. Wild-type (WT) and TLR9 knockout (TLR9KO) mice (C57BL/6 background, n = 5) were pre-immunized intraperitoneally with 1 × 108 formalin-fixed P. gingivalis and boosted once with 1 × 107 formalin-fixed P. gingivalis. Isolated splenocytes and purified B cells from each mouse were cultured with 1 × 108 formalin-fixed P. gingivalis for 48 h. Immunocytochemistry was performed to detect CD45+ IL-10+ cells. Levels of IL-10 expression and secretion in splenocytes and B cells were detected using qRT-PCR and ELISA, respectively. After stimulation with fixed P. gingivalis, the percentage of CD45+ IL-10+ B cells and the level of IL-10 expression were significantly increased (p < 0.01) in splenocytes and purified B cells isolated from WT mice. However, these changes were not observed in splenocytes and purified B cells from TLR9KO mice when the cells were treated with fixed P. gingivalis. The percentage of CD45+ IL-10+ B cells was significantly reduced in splenocytes and purified B cells from TLR9KO mice compared to those from WT mice when challenged with P. gingivalis. IL-10 expression in B cells from TLR9KO mice was significantly decreased compared to those from WT mice at both the mRNA and protein levels. Additionally, P. gingivalis-induced up-regulation of TNF-α mRNA expressions were consistently observed in B cells from both WT and TLR9KO mice. P. gingivalis-induced B10 activation and IL-10 production during adaptive responses by primed B cells requires TLR9 signaling and can be achieved independent of T-cell help.
Keywords: B10 cells; P. gingivalis; TLR-9; adaptive immune response; periodontal disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
