

Blue light promotes ascorbate synthesis by deactivating the PAS/LOV photoreceptor that inhibits GDP-L-galactose phosphorylase
- PMID: 37052931
- PMCID: PMC10291033
- DOI: 10.1093/plcell/koad108
Blue light promotes ascorbate synthesis by deactivating the PAS/LOV photoreceptor that inhibits GDP-L-galactose phosphorylase
Abstract
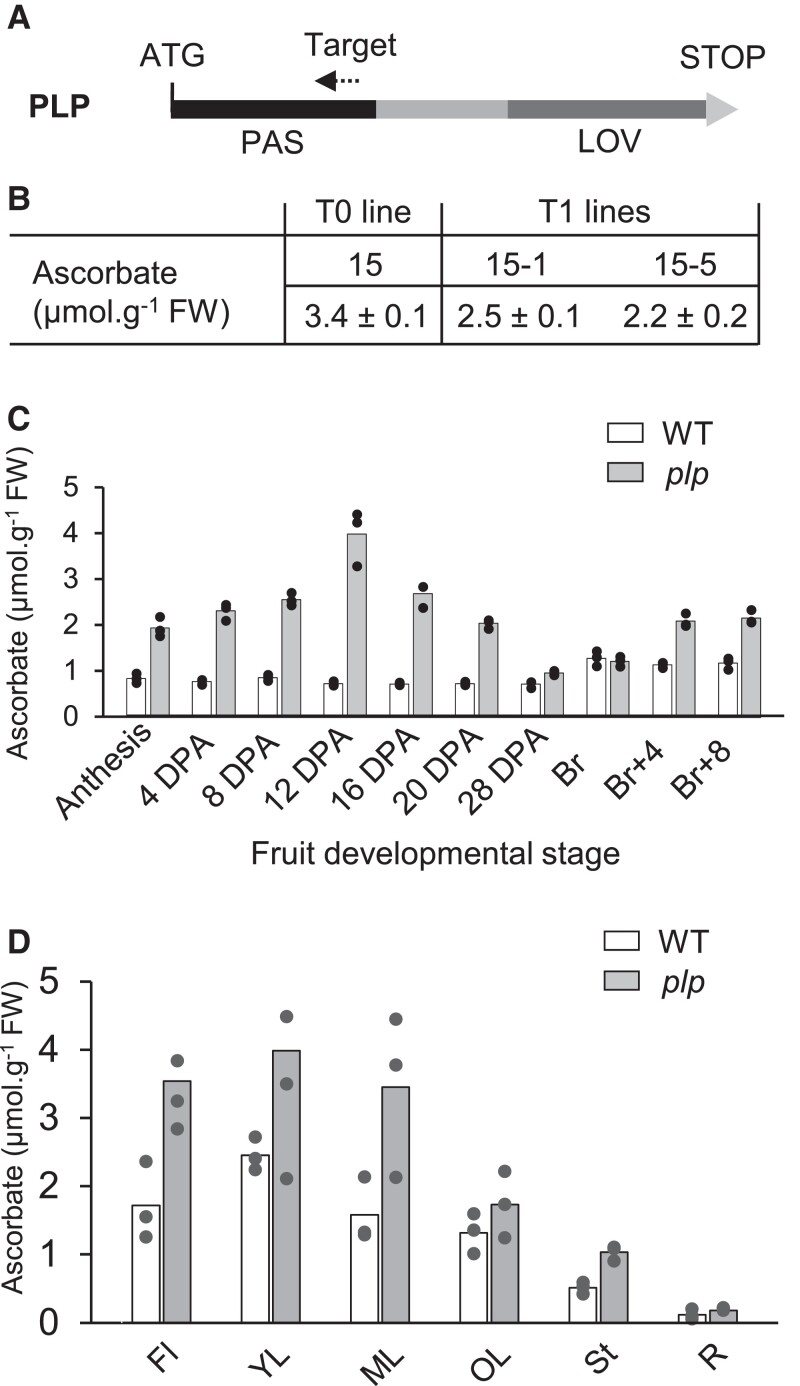
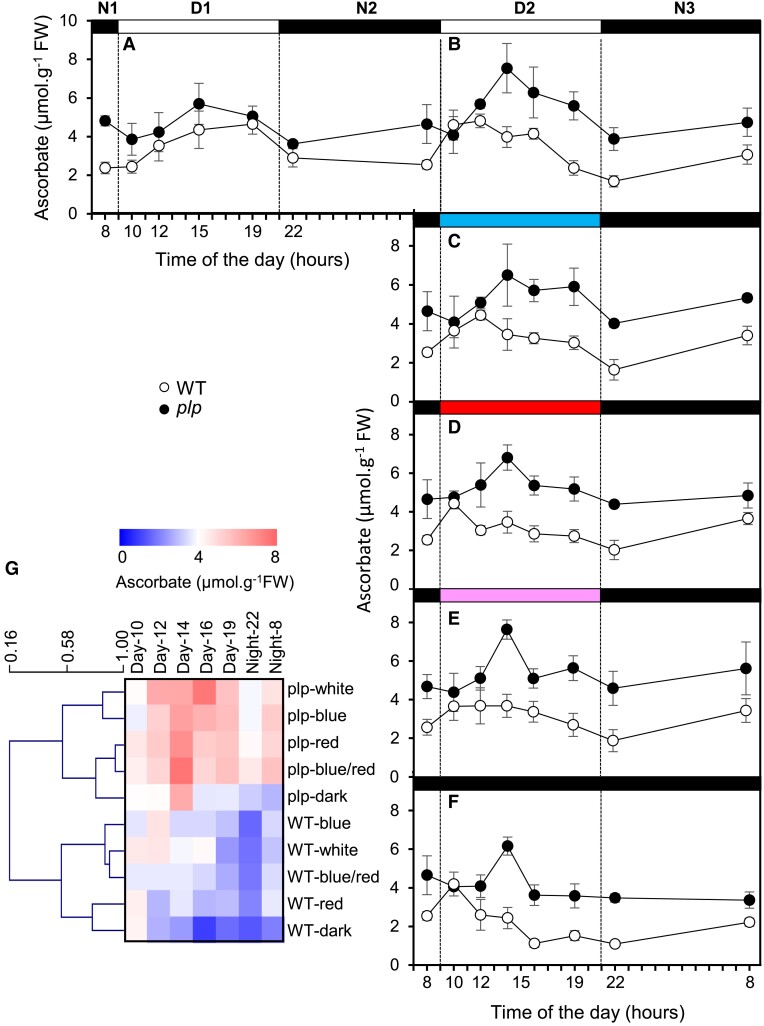
Ascorbate (vitamin C) is an essential antioxidant in fresh fruits and vegetables. To gain insight into the regulation of ascorbate metabolism in plants, we studied mutant tomato plants (Solanum lycopersicum) that produce ascorbate-enriched fruits. The causal mutation, identified by a mapping-by-sequencing strategy, corresponded to a knock-out recessive mutation in a class of photoreceptor named PAS/LOV protein (PLP), which acts as a negative regulator of ascorbate biosynthesis. This trait was confirmed by CRISPR/Cas9 gene editing and further found in all plant organs, including fruit that accumulated 2 to 3 times more ascorbate than in the WT. The functional characterization revealed that PLP interacted with the 2 isoforms of GDP-L-galactose phosphorylase (GGP), known as the controlling step of the L-galactose pathway of ascorbate synthesis. The interaction with GGP occurred in the cytoplasm and the nucleus, but was abolished when PLP was truncated. These results were confirmed by a synthetic approach using an animal cell system, which additionally demonstrated that blue light modulated the PLP-GGP interaction. Assays performed in vitro with heterologously expressed GGP and PLP showed that PLP is a noncompetitive inhibitor of GGP that is inactivated after blue light exposure. This discovery provides a greater understanding of the light-dependent regulation of ascorbate metabolism in plants.
© The Author(s) 2023. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures





Comment in
-
Out of the blue: blue light mediates ascorbate synthesis.Plant Cell. 2023 Jun 26;35(7):2440-2441. doi: 10.1093/plcell/koad109. Plant Cell. 2023. PMID: 37043545 Free PMC article. No abstract available.
-
Elucidating the role of ascorbate in light signaling.Trends Plant Sci. 2023 Sep;28(9):978-980. doi: 10.1016/j.tplants.2023.05.007. Epub 2023 May 27. Trends Plant Sci. 2023. PMID: 37246016
References
-
- Alimohammfadi M, de Silva K, Ballu C, Ali N, Khodakovskaya MV. Reduction of inositol (1,4,5)–trisphosphate affects the overall phosphoinositol pathway and leads to modifications in light signalling and secondary metabolism in tomato plants. J Exp Bot. 2012:63(2):825–835. 10.1093/jxb/err306 - DOI - PMC - PubMed
-
- Bergmeyer HU. Methods of enzymatic analysis. Weinheim: VCH. 1987
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
