

Heterozygous inversion breakpoints suppress meiotic crossovers by altering recombination repair outcomes
- PMID: 37053290
- PMCID: PMC10128924
- DOI: 10.1371/journal.pgen.1010702
Heterozygous inversion breakpoints suppress meiotic crossovers by altering recombination repair outcomes
Abstract
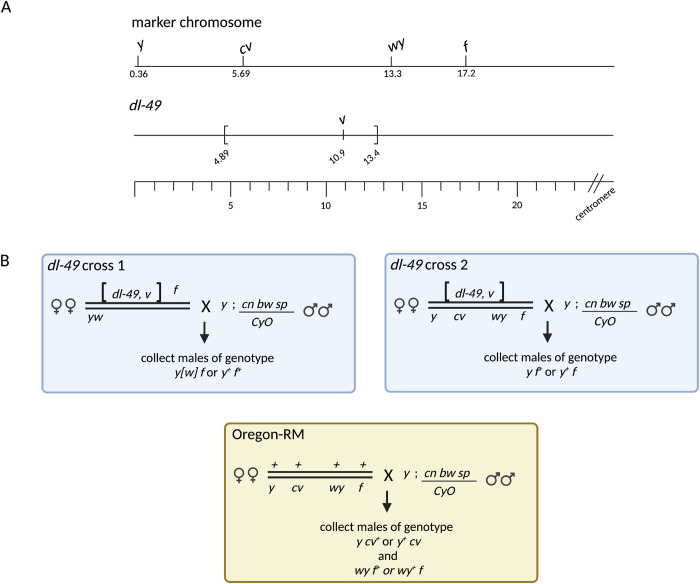
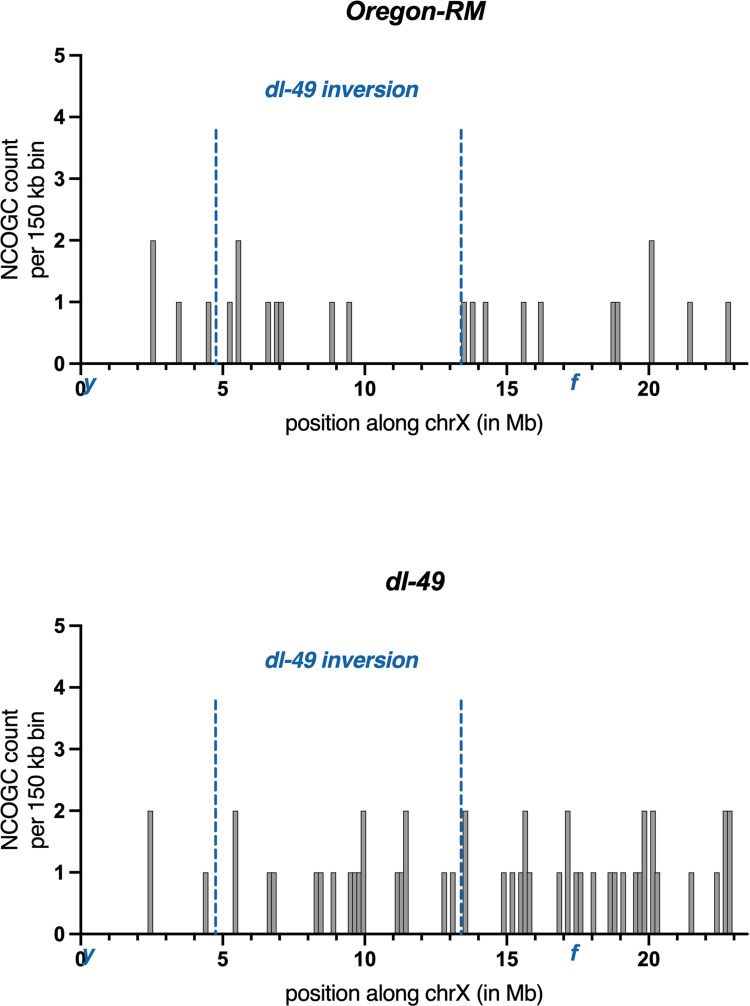
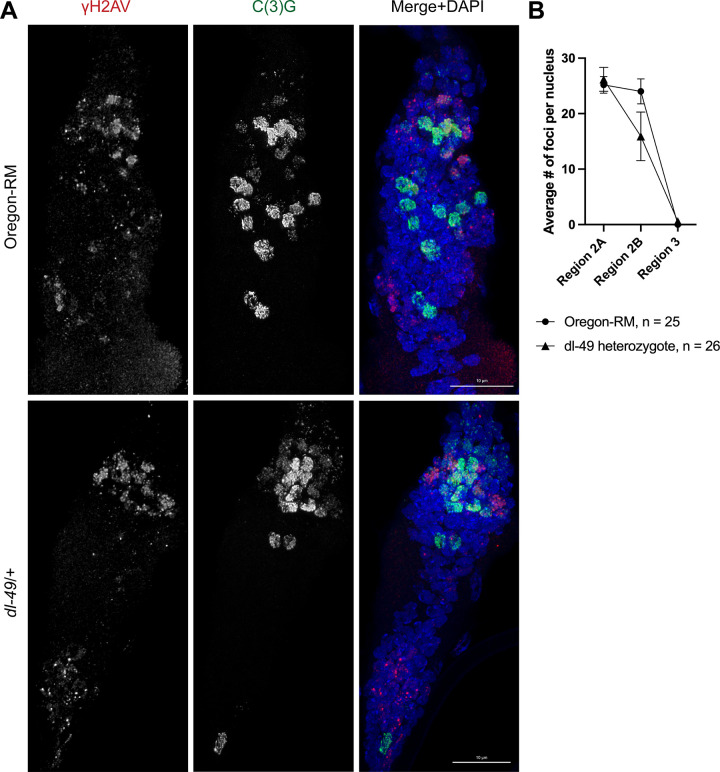
Heterozygous chromosome inversions suppress meiotic crossover (CO) formation within an inversion, potentially because they lead to gross chromosome rearrangements that produce inviable gametes. Interestingly, COs are also severely reduced in regions nearby but outside of inversion breakpoints even though COs in these regions do not result in rearrangements. Our mechanistic understanding of why COs are suppressed outside of inversion breakpoints is limited by a lack of data on the frequency of noncrossover gene conversions (NCOGCs) in these regions. To address this critical gap, we mapped the location and frequency of rare CO and NCOGC events that occurred outside of the dl-49 chrX inversion in D. melanogaster. We created full-sibling wildtype and inversion stocks and recovered COs and NCOGCs in the syntenic regions of both stocks, allowing us to directly compare rates and distributions of recombination events. We show that COs outside of the proximal inversion breakpoint are distributed in a distance-dependent manner, with strongest suppression near the inversion breakpoint. We find that NCOGCs occur evenly throughout the chromosome and, importantly, are not suppressed near inversion breakpoints. We propose a model in which COs are suppressed by inversion breakpoints in a distance-dependent manner through mechanisms that influence DNA double-strand break repair outcome but not double-strand break formation. We suggest that subtle changes in the synaptonemal complex and chromosome pairing might lead to unstable interhomolog interactions during recombination that permits NCOGC formation but not CO formation.
Copyright: © 2023 Li et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: DEM is engaged in a research agreement with Oxford Nanopore Technologies and they have paid for him to travel to speak on their behalf. DEM is also on the scientific advisory board of Oxford Nanopore Technologies.
Figures


References
-
- Sturtevant AH. A crossover reducer in Drosophila melanogaster due to inversion of a section of the third chromosome. Biologisches Zentralblatt. 1926;46(12):697–702.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
