

Bone-derived C-terminal FGF23 cleaved peptides increase iron availability in acute inflammation
- PMID: 37053547
- PMCID: PMC10356820
- DOI: 10.1182/blood.2022018475
Bone-derived C-terminal FGF23 cleaved peptides increase iron availability in acute inflammation
Abstract
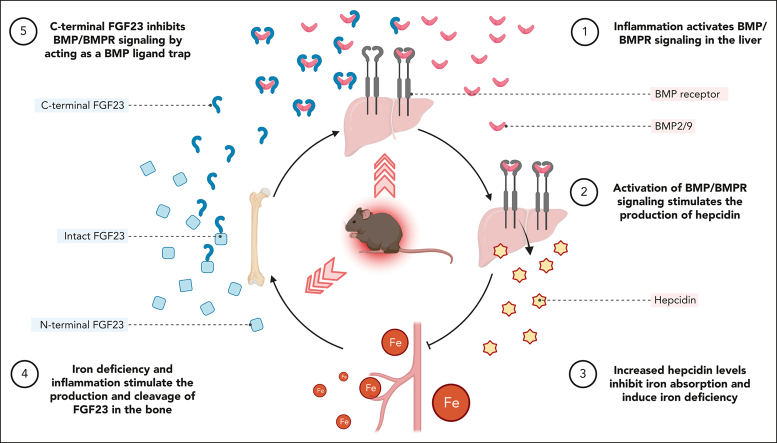
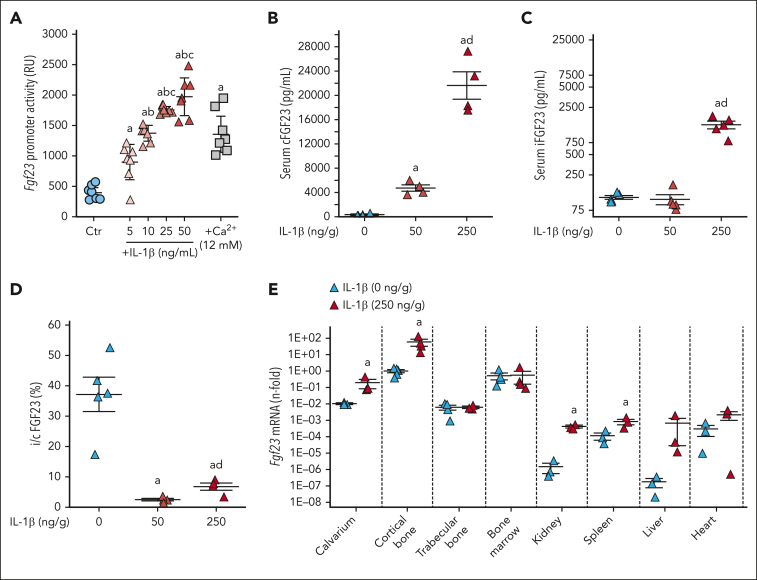
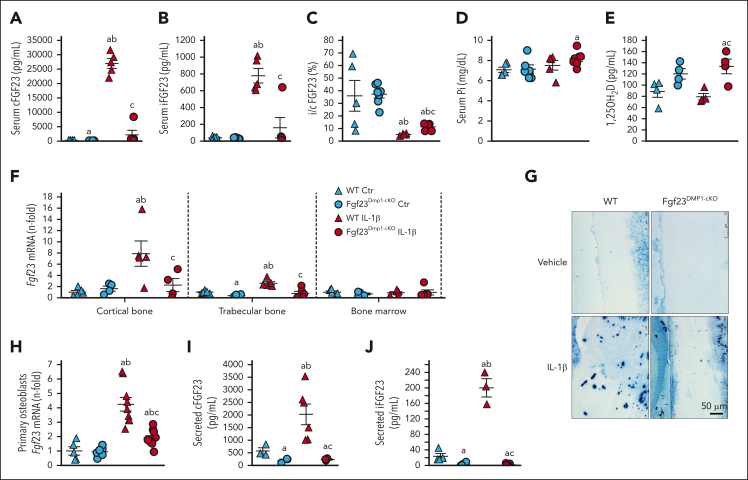
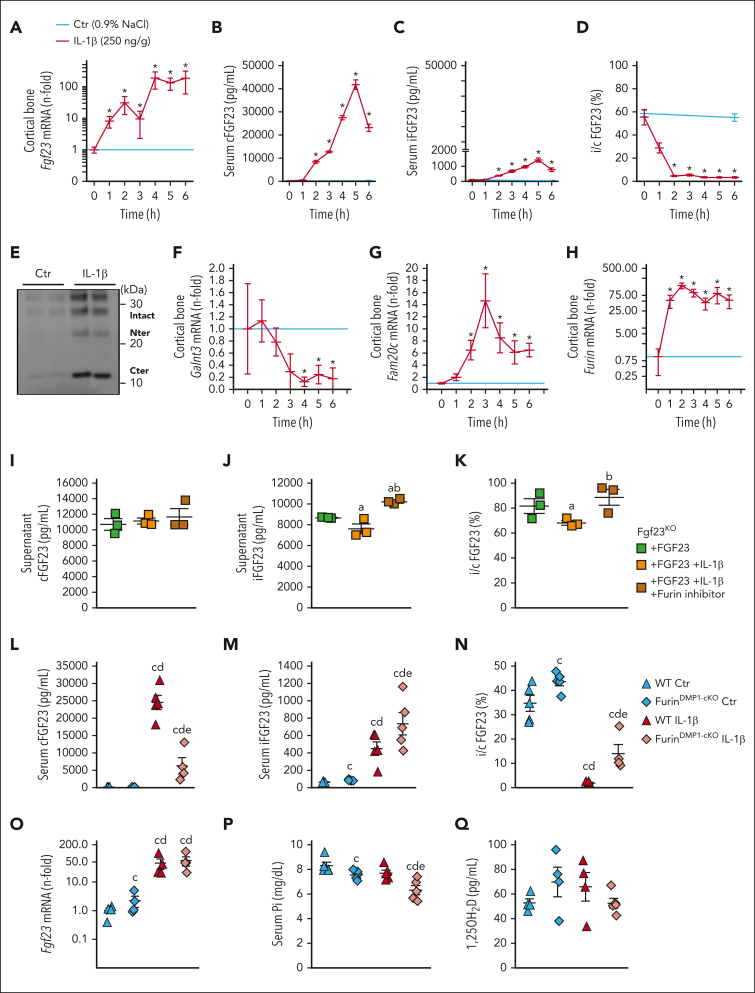
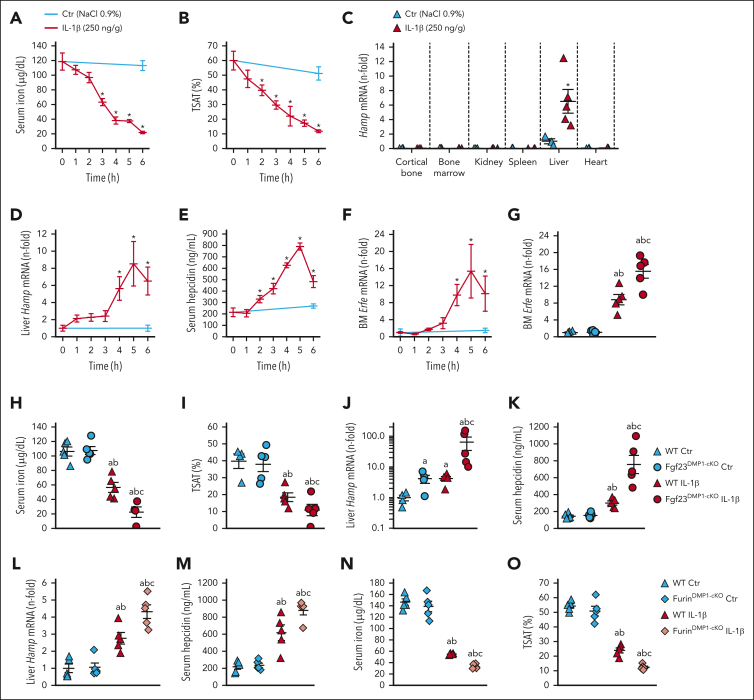
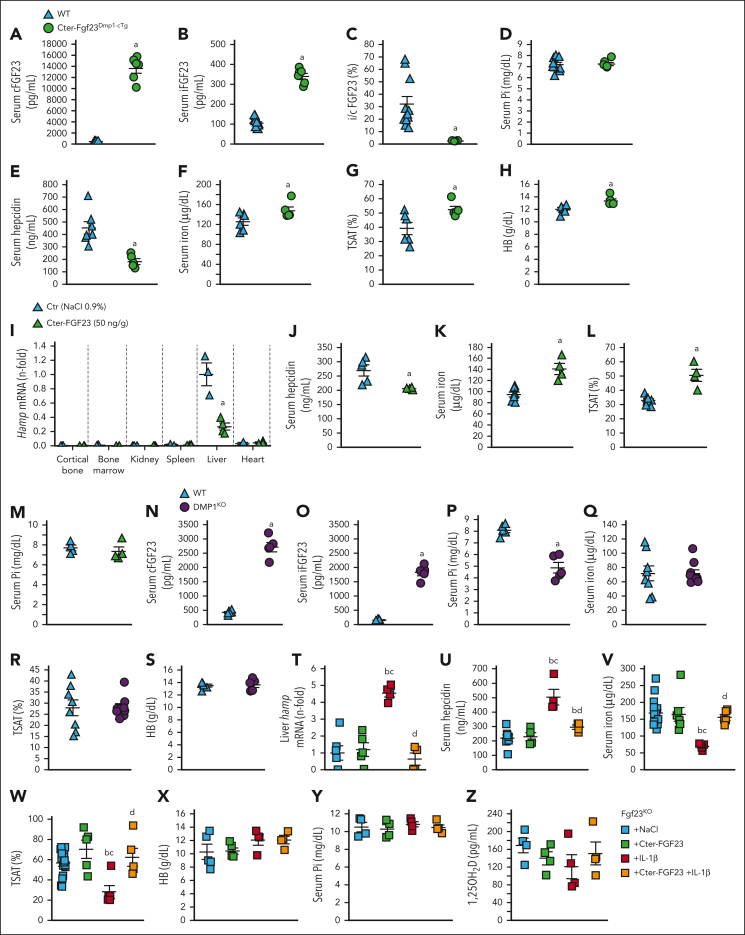
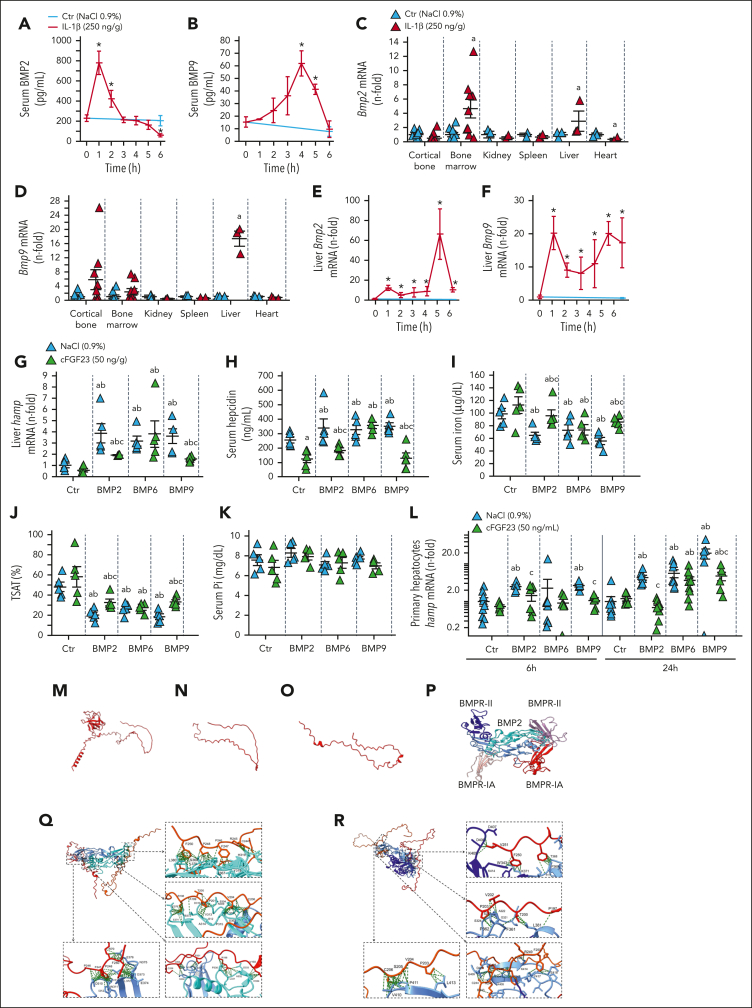
Inflammation leads to functional iron deficiency by increasing the expression of the hepatic iron regulatory peptide hepcidin. Inflammation also stimulates fibroblast growth factor 23 (FGF23) production by increasing both Fgf23 transcription and FGF23 cleavage, which paradoxically leads to excess in C-terminal FGF23 peptides (Cter-FGF23), rather than intact FGF23 (iFGF23) hormone. We determined that the major source of Cter-FGF23 is osteocytes and investigated whether Cter-FGF23 peptides play a direct role in the regulation of hepcidin and iron metabolism in response to acute inflammation. Mice harboring an osteocyte-specific deletion of Fgf23 showed a ∼90% reduction in Cter-FGF23 levels during acute inflammation. Reduction in Cter-FGF23 led to a further decrease in circulating iron in inflamed mice owing to excessive hepcidin production. We observed similar results in mice showing impaired FGF23 cleavage owing to osteocyte-specific deletion of Furin. We next showed that Cter-FGF23 peptides bind members of the bone morphogenetic protein (BMP) family, BMP2 and BMP9, which are established inducers of hepcidin. Coadministration of Cter-FGF23 and BMP2 or BMP9 prevented the increase in Hamp messenger RNA and circulating hepcidin levels induced by BMP2/9, resulting in normal serum iron levels. Finally, injection of Cter-FGF23 in inflamed Fgf23KO mice and genetic overexpression of Cter-Fgf23 in wild type mice also resulted in lower hepcidin and higher circulating iron levels. In conclusion, during inflammation, bone is the major source of Cter-FGF23 secretion, and independently of iFGF23, Cter-FGF23 reduces BMP-induced hepcidin secretion in the liver.
© 2023 by The American Society of Hematology. Licensed under Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0), permitting only noncommercial, nonderivative use with attribution. All other rights reserved.
Conflict of interest statement
Conflict-of-interest disclosure: V.D. received research funding from Akebia and has received research funding from Vifor Pharma and consulting honoraria from Keryx Biopharmaceuticals, Vifor Pharma, Luitpold, and Amgen, outside of the submitted work. The remaining authors declare no competing financial interests.
Figures
Comment in
-
Matryoshka hormones.Blood. 2023 Jul 6;142(1):7-9. doi: 10.1182/blood.2023020770. Blood. 2023. PMID: 37410509 No abstract available.
References
-
- Liu S, Zhou J, Tang W, Jiang X, Rowe DW, Quarles LD. Pathogenic role of Fgf23 in Hyp mice. Am J Physiol Endocrinol Metab. 2006;291(1):E38–E49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK101730/DK/NIDDK NIH HHS/United States
- IS1 BX004813/BX/BLRD VA/United States
- R01 DK132657/DK/NIDDK NIH HHS/United States
- IS1 BX003589/BX/BLRD VA/United States
- R01 DK124220/DK/NIDDK NIH HHS/United States
- K24 HL150235/HL/NHLBI NIH HHS/United States
- R01 DK122259/DK/NIDDK NIH HHS/United States
- IK6 BX004835/BX/BLRD VA/United States
- R01 DK131046/DK/NIDDK NIH HHS/United States
- I01 BX005851/BX/BLRD VA/United States
- P30 AR066262/AR/NIAMS NIH HHS/United States
- IS1 BX005042/BX/BLRD VA/United States
- R01 HL150729/HL/NHLBI NIH HHS/United States
- R01 DK132342/DK/NIDDK NIH HHS/United States
- RF1 AG075742/AG/NIA NIH HHS/United States
- R01 HL148012/HL/NHLBI NIH HHS/United States
- R01 DK102815/DK/NIDDK NIH HHS/United States
- R01 HL169507/HL/NHLBI NIH HHS/United States
- R01 DK114158/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
