

Bi-allelic variants in INTS11 are associated with a complex neurological disorder
- PMID: 37054711
- PMCID: PMC10183469
- DOI: 10.1016/j.ajhg.2023.03.012
Bi-allelic variants in INTS11 are associated with a complex neurological disorder
Abstract
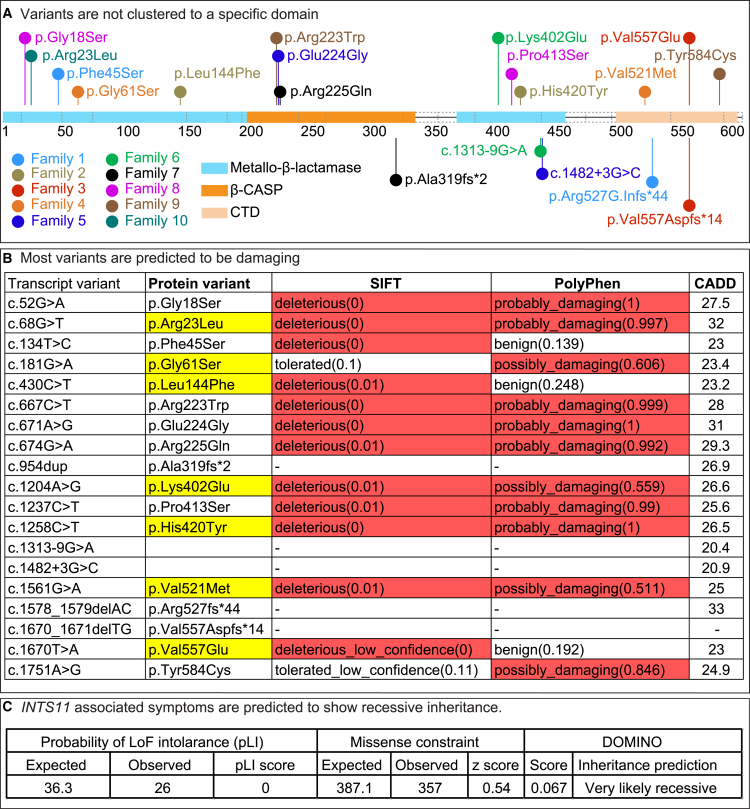
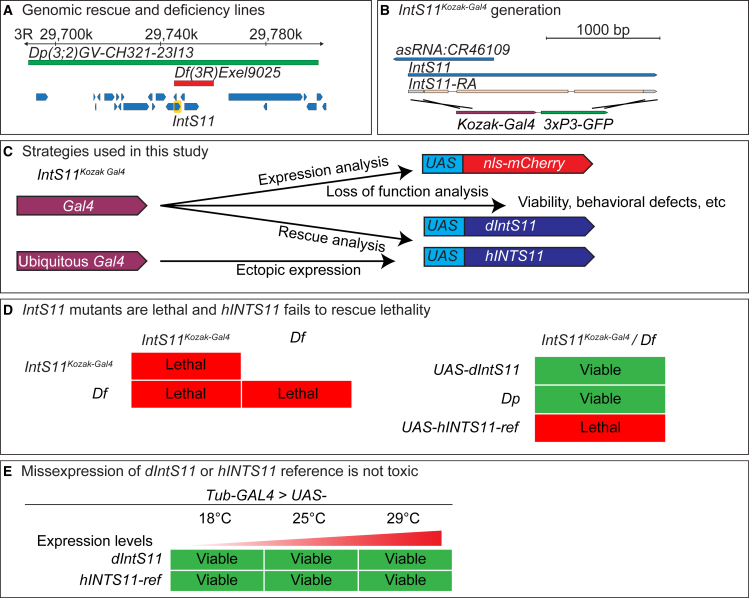
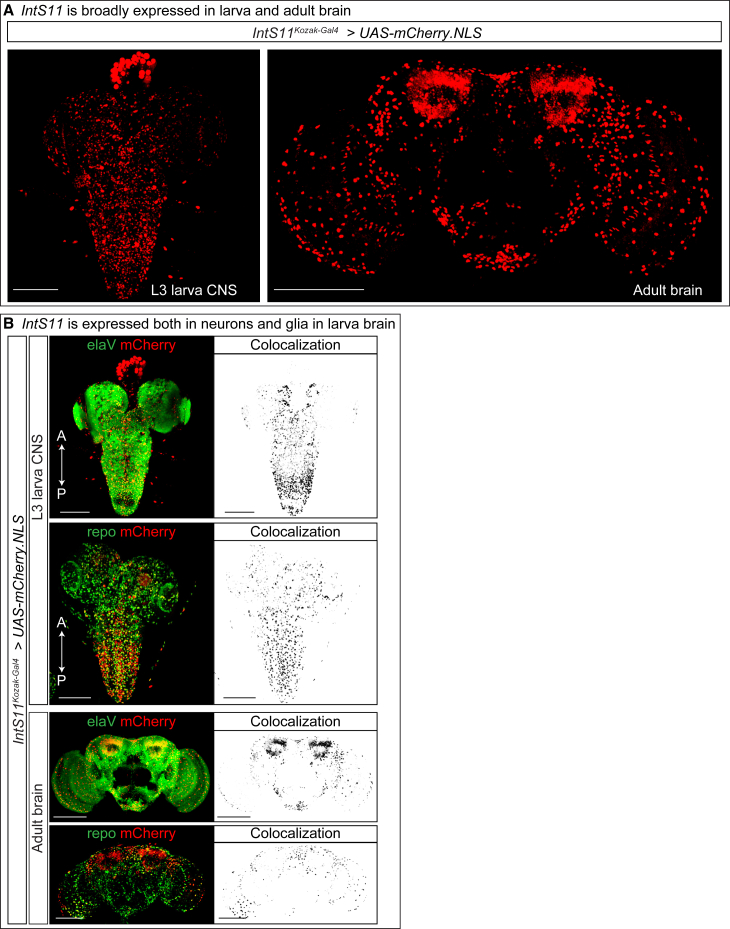
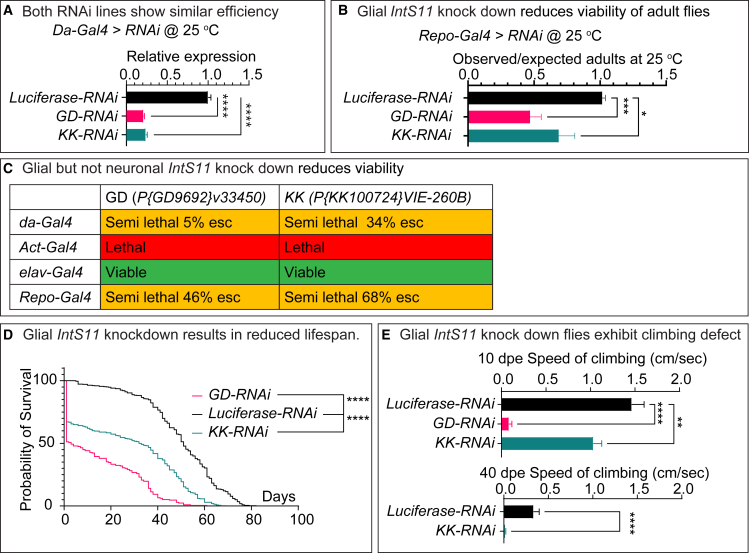
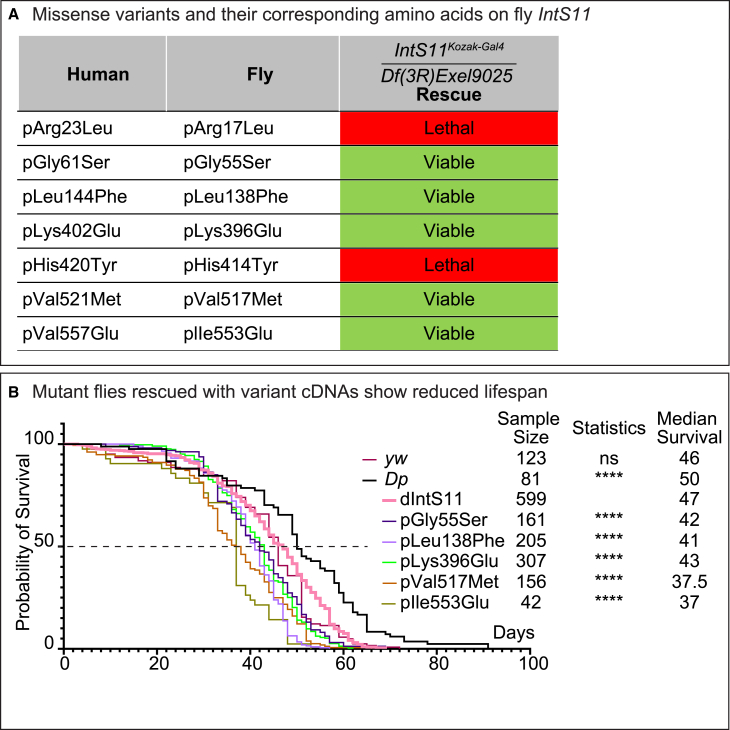
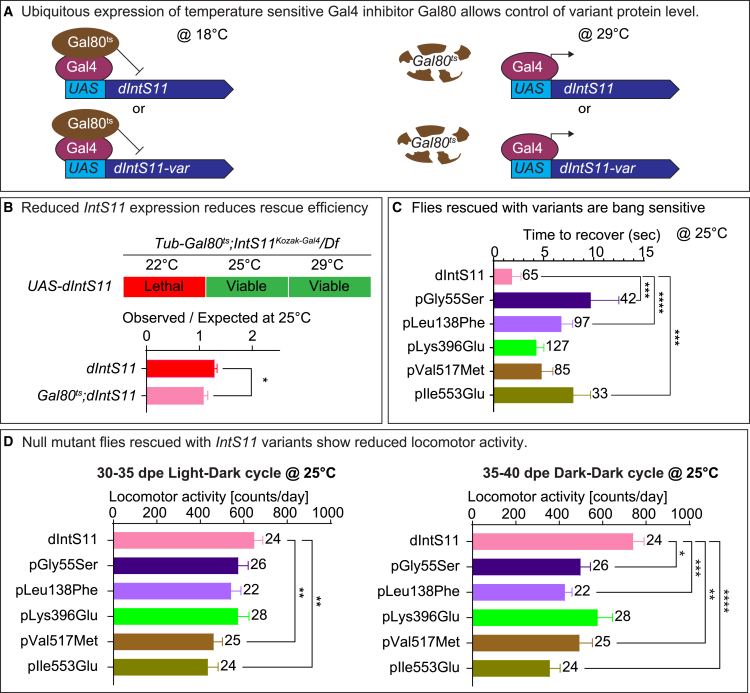
The Integrator complex is a multi-subunit protein complex that regulates the processing of nascent RNAs transcribed by RNA polymerase II (RNAPII), including small nuclear RNAs, enhancer RNAs, telomeric RNAs, viral RNAs, and protein-coding mRNAs. Integrator subunit 11 (INTS11) is the catalytic subunit that cleaves nascent RNAs, but, to date, mutations in this subunit have not been linked to human disease. Here, we describe 15 individuals from 10 unrelated families with bi-allelic variants in INTS11 who present with global developmental and language delay, intellectual disability, impaired motor development, and brain atrophy. Consistent with human observations, we find that the fly ortholog of INTS11, dIntS11, is essential and expressed in the central nervous systems in a subset of neurons and most glia in larval and adult stages. Using Drosophila as a model, we investigated the effect of seven variants. We found that two (p.Arg17Leu and p.His414Tyr) fail to rescue the lethality of null mutants, indicating that they are strong loss-of-function variants. Furthermore, we found that five variants (p.Gly55Ser, p.Leu138Phe, p.Lys396Glu, p.Val517Met, and p.Ile553Glu) rescue lethality but cause a shortened lifespan and bang sensitivity and affect locomotor activity, indicating that they are partial loss-of-function variants. Altogether, our results provide compelling evidence that integrity of the Integrator RNA endonuclease is critical for brain development.
Keywords: CPSF3L; Drosophila; INTS11; brain atrophy; dIntS11; delayed language development; developmental delay; impaired motor development; intellectual disability.
Copyright © 2023 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Nojima T., Proudfoot N.J. Mechanisms of lncRNA biogenesis as revealed by nascent transcriptomics. Nat. Rev. Mol. Cell Biol. 2022;23:389–406. - PubMed
-
- Baillat D., Hakimi M.A., Näär A.M., Shilatifard A., Cooch N., Shiekhattar R. Integrator, a multiprotein mediator of small nuclear RNA processing, associates with the C-terminal repeat of RNA polymerase II. Cell. 2005;123:265–276. - PubMed
-
- Jurica M.S., Moore M.J. Pre-mRNA splicing: awash in a sea of proteins. Mol. Cell. 2003;12:5–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- UM1 HG008900/HG/NHGRI NIH HHS/United States
- P50 HD105351/HD/NICHD NIH HHS/United States
- Z01 OD022005/ImNIH/Intramural NIH HHS/United States
- R24 OD031447/OD/NIH HHS/United States
- R01 HG009141/HG/NHGRI NIH HHS/United States
- U01 NS134356/NS/NINDS NIH HHS/United States
- P30 HD024064/HD/NICHD NIH HHS/United States
- R01 AG073260/AG/NIA NIH HHS/United States
- R01 NS035129/NS/NINDS NIH HHS/United States
- R01 GM134539/GM/NIGMS NIH HHS/United States
- U54 NS093793/NS/NINDS NIH HHS/United States
- R24 OD022005/OD/NIH HHS/United States
- NIHR-RP-2016-07-019/DH_/Department of Health/United Kingdom
- P50 HD103555/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
