

Cross-Platform Synaptic Network Analysis of Human Entorhinal Cortex Identifies TWF2 as a Modulator of Dendritic Spine Length
- PMID: 37055180
- PMCID: PMC10198456
- DOI: 10.1523/JNEUROSCI.2102-22.2023
Cross-Platform Synaptic Network Analysis of Human Entorhinal Cortex Identifies TWF2 as a Modulator of Dendritic Spine Length
Abstract
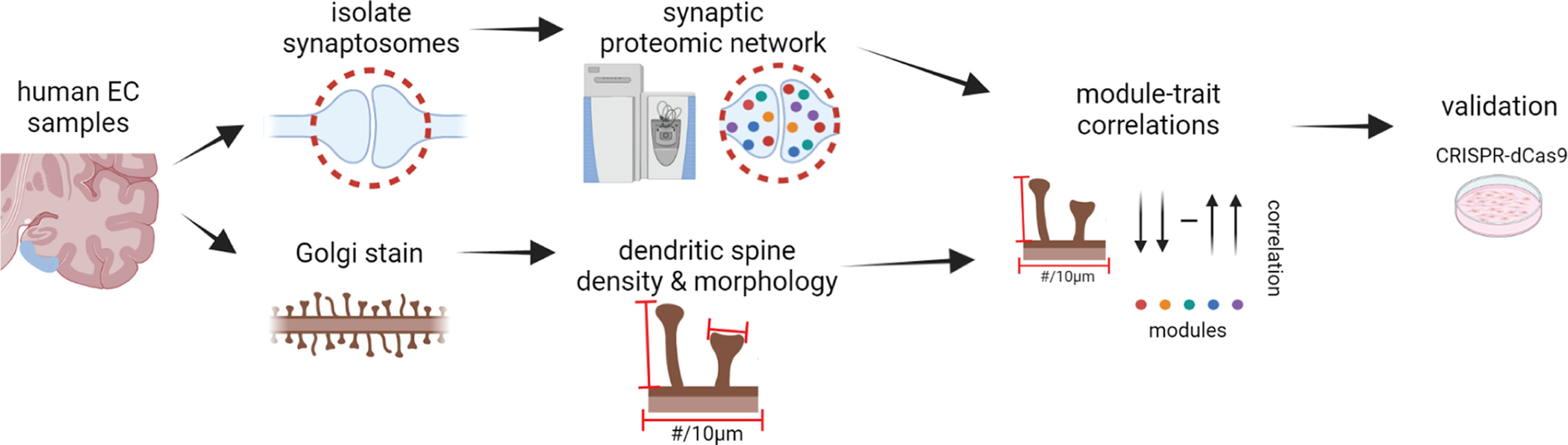
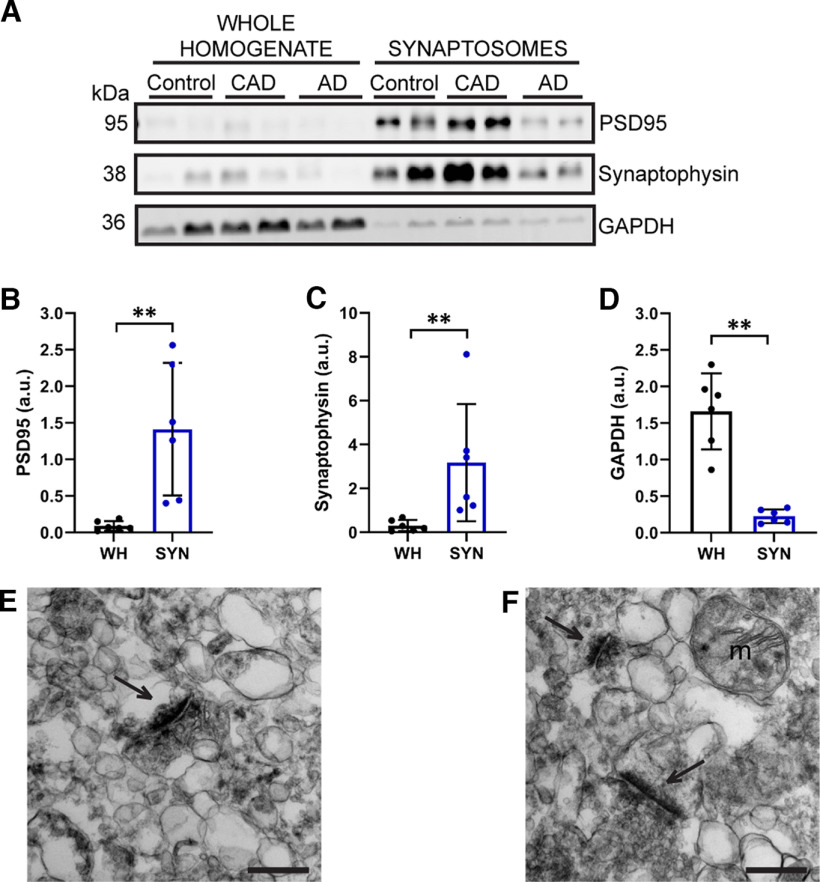
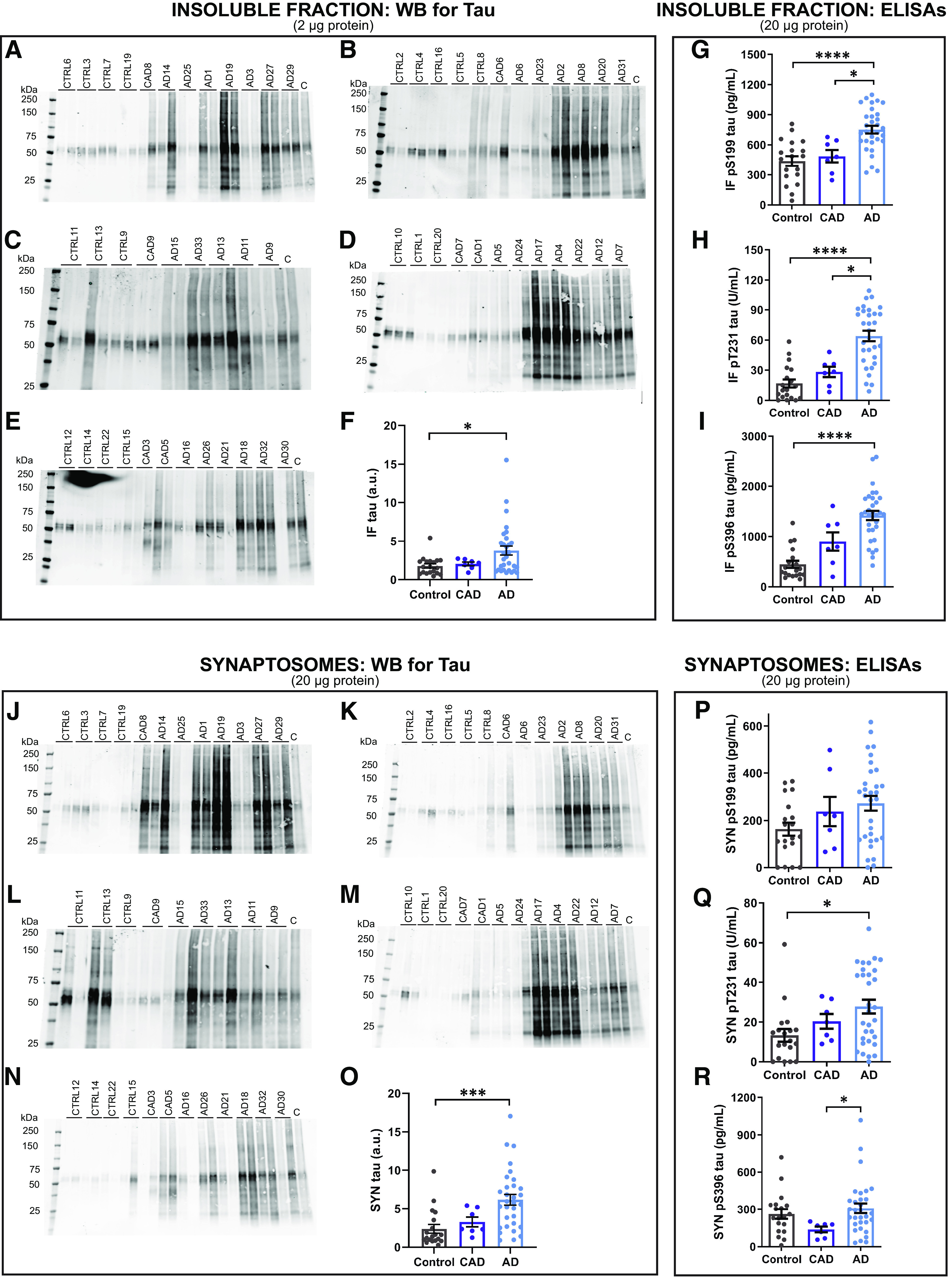
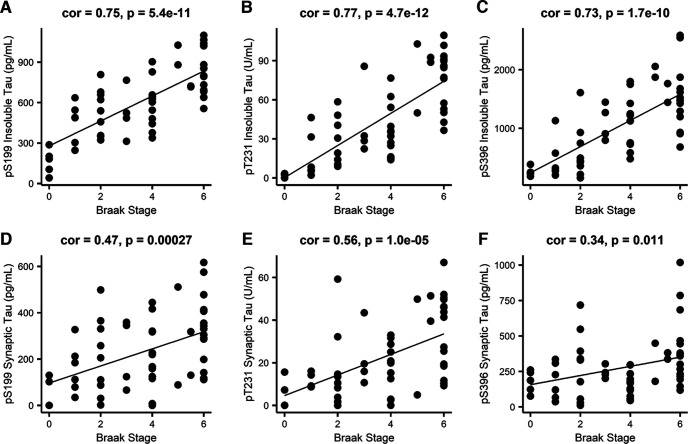
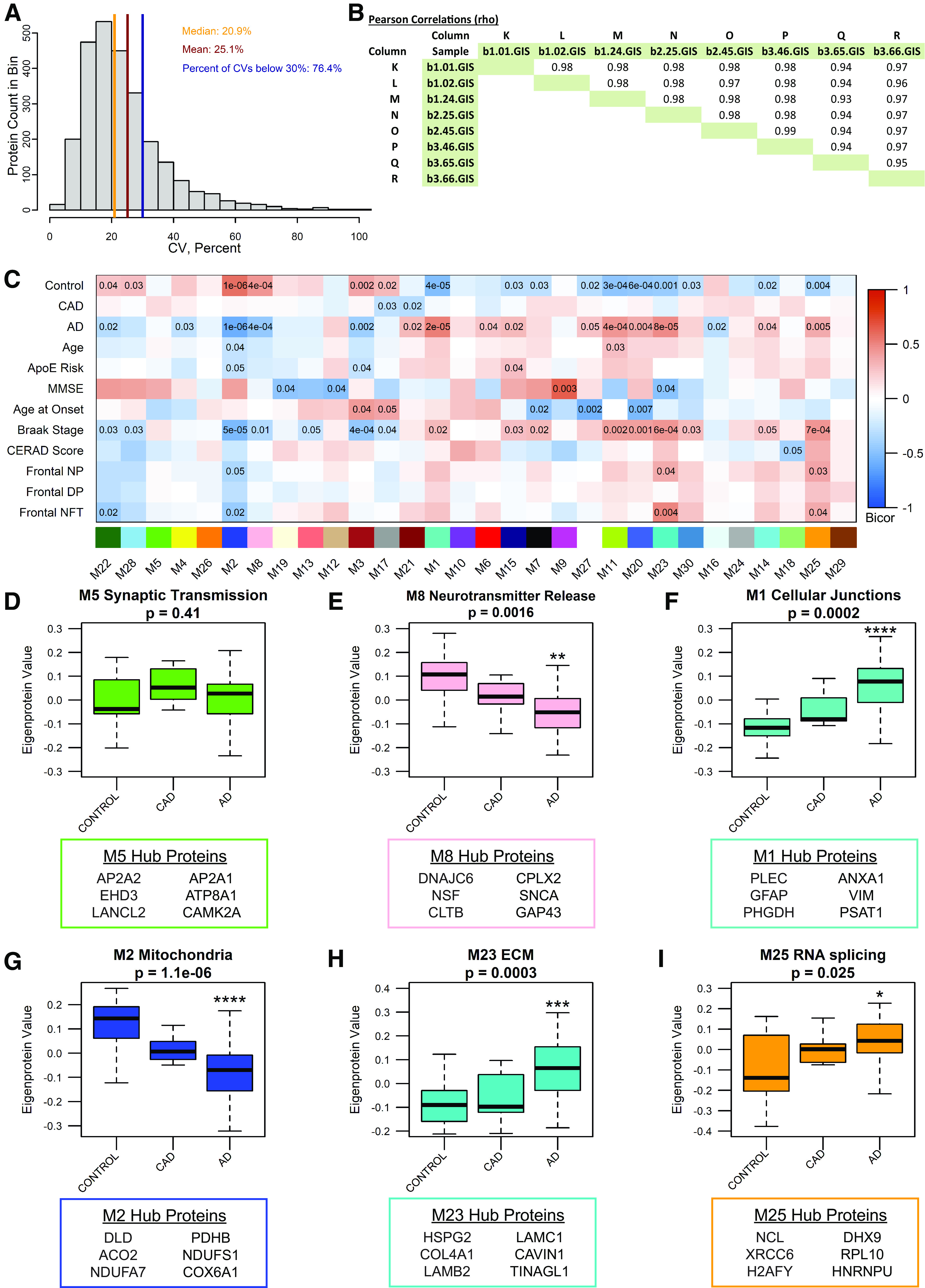
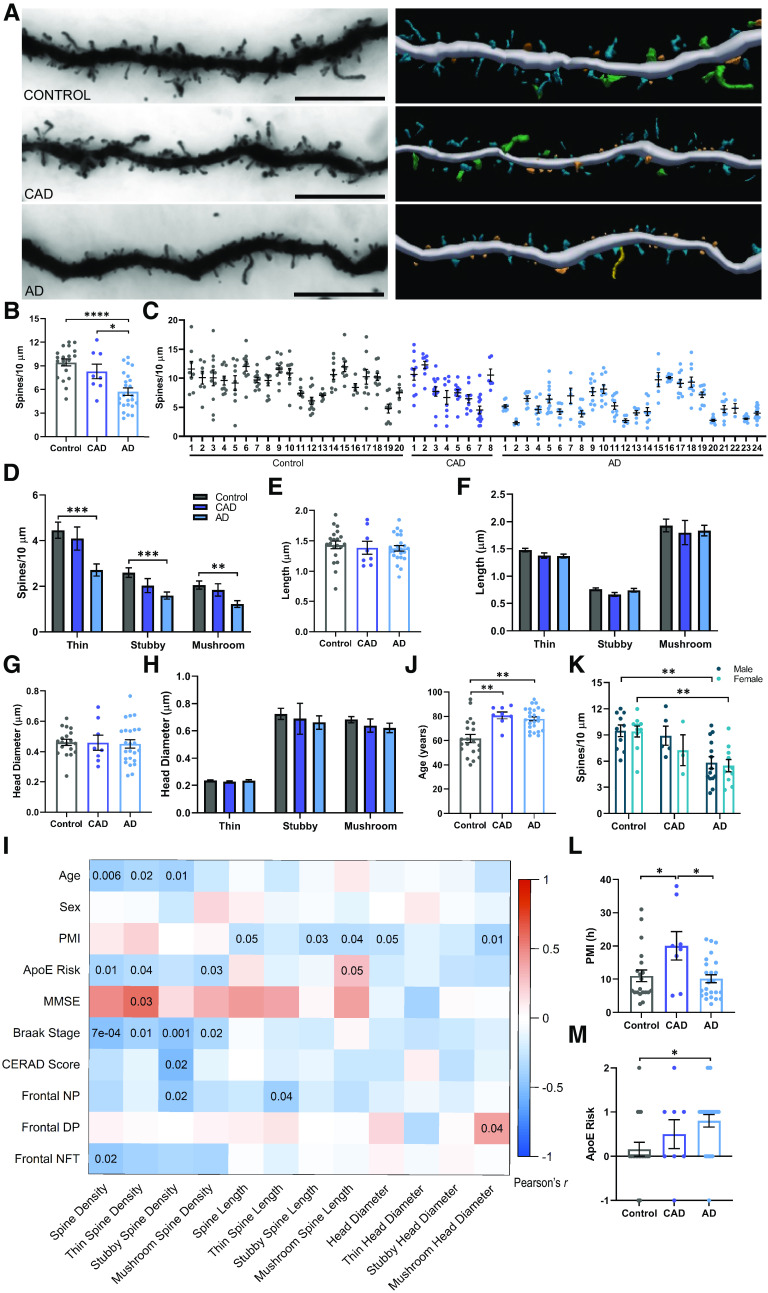
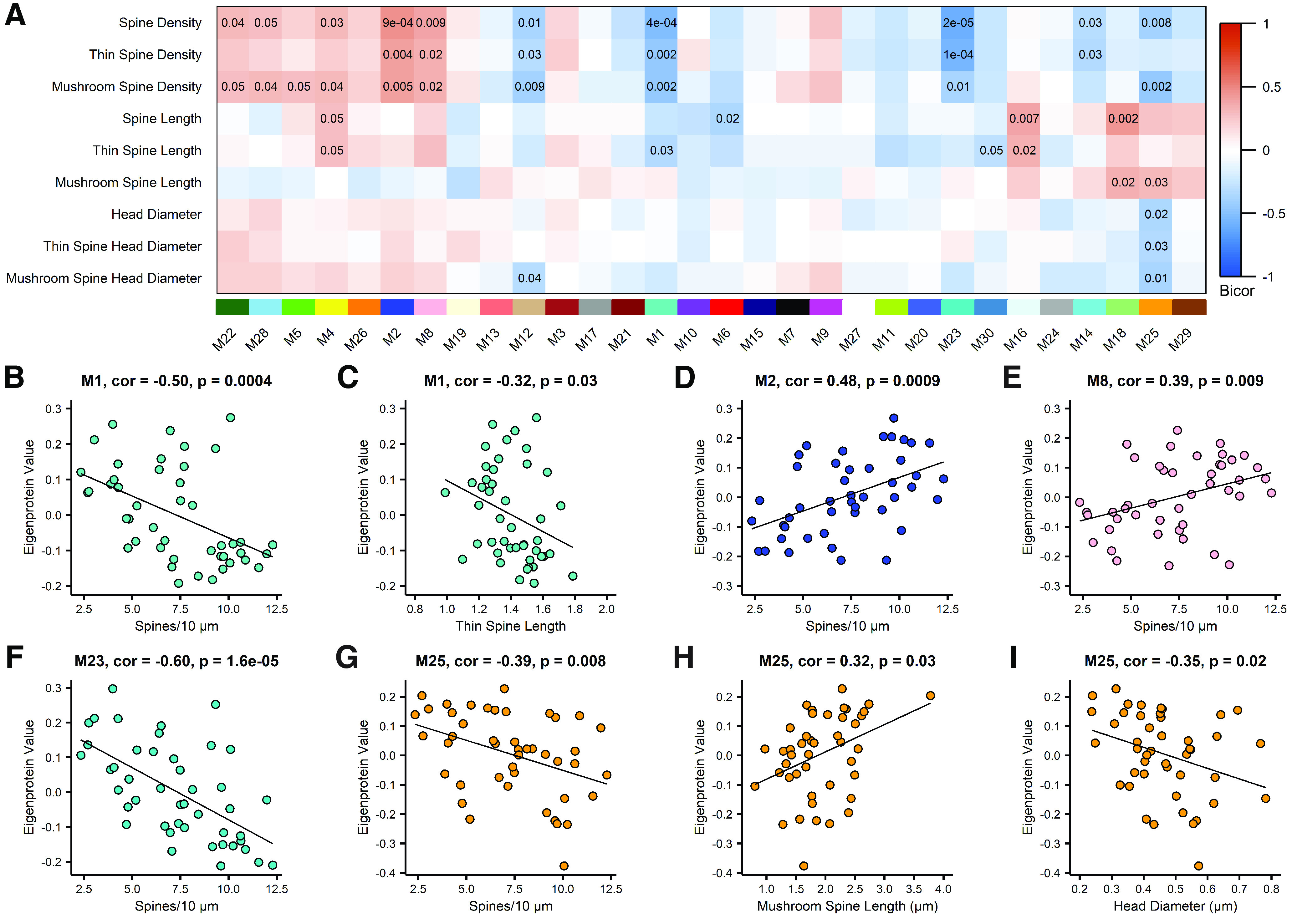
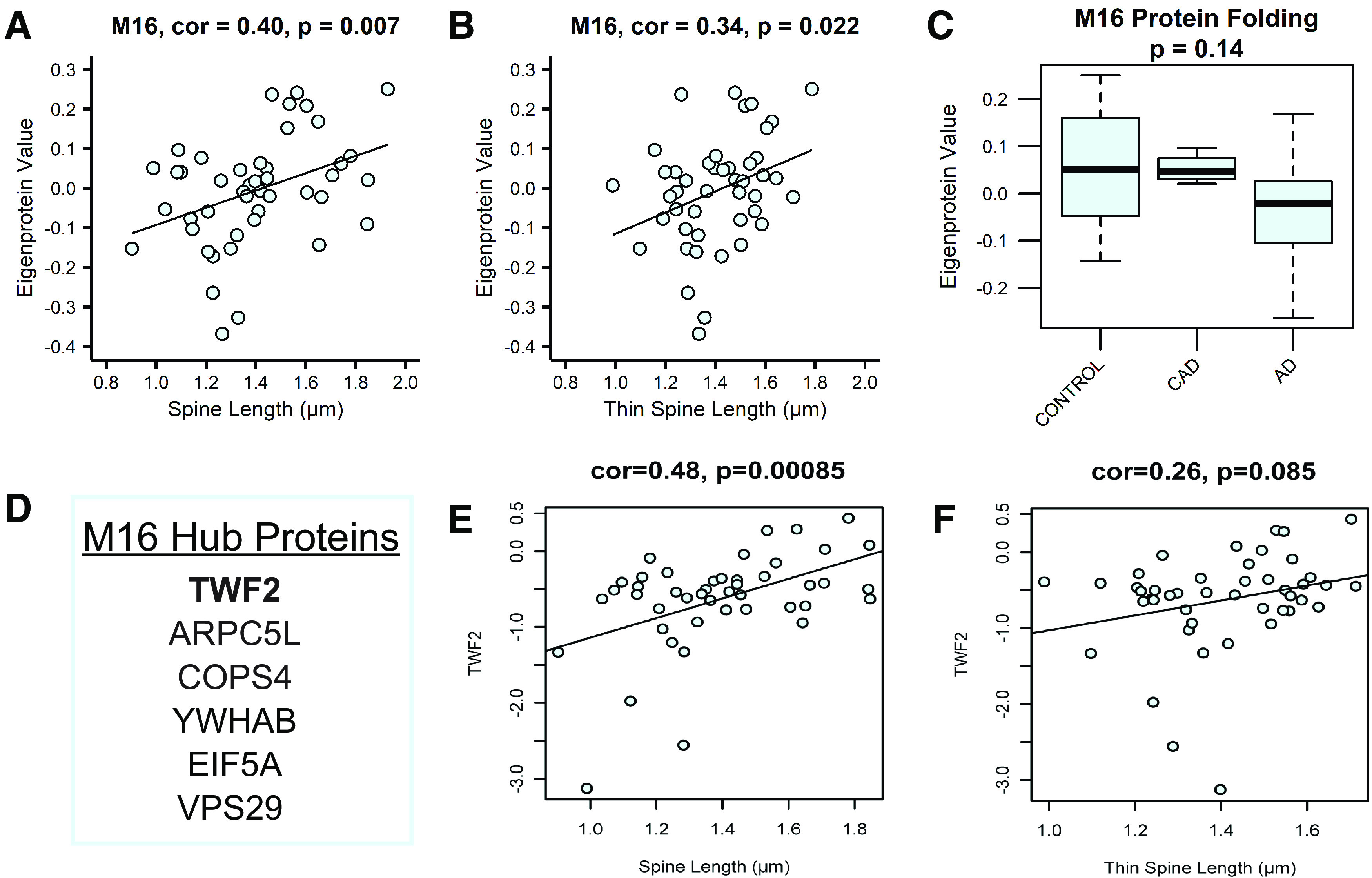
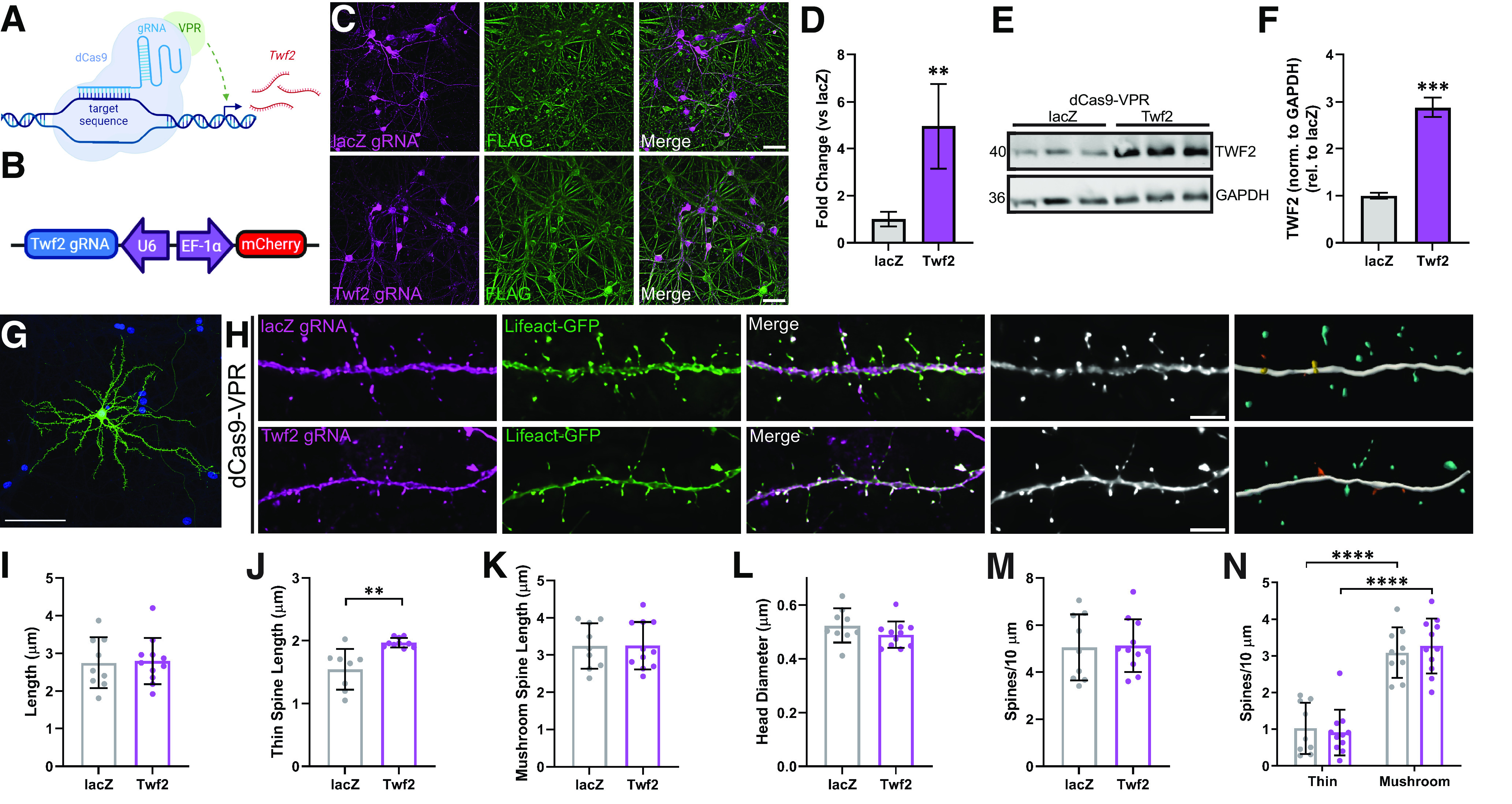
Proteomic studies using postmortem human brain tissue samples have yielded robust assessments of the aging and neurodegenerative disease(s) proteomes. While these analyses provide lists of molecular alterations in human conditions, like Alzheimer's disease (AD), identifying individual proteins that affect biological processes remains a challenge. To complicate matters, protein targets may be highly understudied and have limited information on their function. To address these hurdles, we sought to establish a blueprint to aid selection and functional validation of targets from proteomic datasets. A cross-platform pipeline was engineered to focus on synaptic processes in the entorhinal cortex (EC) of human patients, including controls, preclinical AD, and AD cases. Label-free quantification mass spectrometry (MS) data (n = 2260 proteins) was generated on synaptosome fractionated tissue from Brodmann area 28 (BA28; n = 58 samples). In parallel, dendritic spine density and morphology was measured in the same individuals. Weighted gene co-expression network analysis was used to construct a network of protein co-expression modules that were correlated with dendritic spine metrics. Module-trait correlations were used to guide unbiased selection of Twinfilin-2 (TWF2), which was the top hub protein of a module that positively correlated with thin spine length. Using CRISPR-dCas9 activation strategies, we demonstrated that boosting endogenous TWF2 protein levels in primary hippocampal neurons increased thin spine length, thus providing experimental validation for the human network analysis. Collectively, this study describes alterations in dendritic spine density and morphology as well as synaptic proteins and phosphorylated tau from the entorhinal cortex of preclinical and advanced stage AD patients.SIGNIFICANCE STATEMENT Proteomic studies can yield vast lists of molecules that are altered under various experimental or disease conditions. Here, we provide a blueprint to facilitate mechanistic validation of protein targets from human brain proteomic datasets. We conducted a proteomic analysis of human entorhinal cortex (EC) samples spanning cognitively normal and Alzheimer's disease (AD) cases with a comparison of dendritic spine morphology in the same samples. Network integration of proteomics with dendritic spine measurements allowed for unbiased discovery of Twinfilin-2 (TWF2) as a regulator of dendritic spine length. A proof-of-concept experiment in cultured neurons demonstrated that altering Twinfilin-2 protein level induced corresponding changes in dendritic spine length, thus providing experimental validation for the computational framework.
Keywords: Alzheimer's disease; dendritic spines; entorhinal cortex; proteomics; synapse; systems biology.
Copyright © 2023 Walker et al.
Figures
Similar articles
-
Integrated Proteomics to Understand the Role of Neuritin (NRN1) as a Mediator of Cognitive Resilience to Alzheimer's Disease.Mol Cell Proteomics. 2023 May;22(5):100542. doi: 10.1016/j.mcpro.2023.100542. Epub 2023 Apr 5. Mol Cell Proteomics. 2023. PMID: 37024090 Free PMC article.
-
Proteomic and Transcriptomic Analyses Reveal Pathological Changes in the Entorhinal Cortex Region that Correlate Well with Dysregulation of Ion Transport in Patients with Alzheimer's Disease.Mol Neurobiol. 2021 Aug;58(8):4007-4027. doi: 10.1007/s12035-021-02356-3. Epub 2021 Apr 27. Mol Neurobiol. 2021. PMID: 33904022
-
3D Electron Microscopy Study of Synaptic Organization of the Normal Human Transentorhinal Cortex and Its Possible Alterations in Alzheimer's Disease.eNeuro. 2019 Jul 10;6(4):ENEURO.0140-19.2019. doi: 10.1523/ENEURO.0140-19.2019. Print 2019 Jul/Aug. eNeuro. 2019. PMID: 31217195 Free PMC article.
-
Amyloid beta: a putative intra-spinal microtubule-depolymerizer to induce synapse-loss or dentritic spine shortening in Alzheimer's disease.Ital J Anat Embryol. 2009 Apr-Sep;114(2-3):109-20. Ital J Anat Embryol. 2009. PMID: 20198823 Review.
-
Network activity changes in the pathophysiology of Alzheimer's disease: the role of aging and early entorhinal cortex dysfunction.Metab Brain Dis. 2022 Feb;37(2):289-298. doi: 10.1007/s11011-021-00848-8. Epub 2021 Sep 30. Metab Brain Dis. 2022. PMID: 34591222 Review.
Cited by
-
Sequence of episodic memory-related behavioral and brain-imaging abnormalities in type 2 diabetes.Nutr Diabetes. 2025 Feb 1;15(1):1. doi: 10.1038/s41387-025-00359-w. Nutr Diabetes. 2025. PMID: 39893169 Free PMC article. Clinical Trial.
-
Cognitive integrity in Non-Demented Individuals with Alzheimer's Neuropathology is associated with preservation and remodeling of dendritic spines.Alzheimers Dement. 2024 Jul;20(7):4677-4691. doi: 10.1002/alz.13900. Epub 2024 Jun 3. Alzheimers Dement. 2024. PMID: 38829680 Free PMC article.
-
CoPPIs algorithm: a tool to unravel protein cooperative strategies in pathophysiological conditions.Brief Bioinform. 2025 Mar 4;26(2):bbaf146. doi: 10.1093/bib/bbaf146. Brief Bioinform. 2025. PMID: 40194557 Free PMC article.
-
Dendritic spine head diameter predicts episodic memory performance in older adults.Sci Adv. 2024 Aug 9;10(32):eadn5181. doi: 10.1126/sciadv.adn5181. Epub 2024 Aug 7. Sci Adv. 2024. PMID: 39110801 Free PMC article.
-
Integration across biophysical scales identifies molecular and cellular correlates of person-to-person variability in human brain connectivity.Nat Neurosci. 2024 Nov;27(11):2240-2252. doi: 10.1038/s41593-024-01788-z. Epub 2024 Oct 31. Nat Neurosci. 2024. PMID: 39482360 Free PMC article.
References
-
- Bilousova T, Miller CA, Poon WW, Vinters HV, Corrada M, Kawas C, Hayden EY, Teplow DB, Glabe C, Albay R 3rd, Cole GM, Teng E, Gylys KH (2016) Synaptic amyloid-β oligomers precede p-Tau and differentiate high pathology control cases. Am J Pathol 186:185–198. 10.1016/j.ajpath.2015.09.018 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials