

Stable endocytic structures navigate the complex pellicle of apicomplexan parasites
- PMID: 37061511
- PMCID: PMC10105704
- DOI: 10.1038/s41467-023-37431-x
Stable endocytic structures navigate the complex pellicle of apicomplexan parasites
Abstract
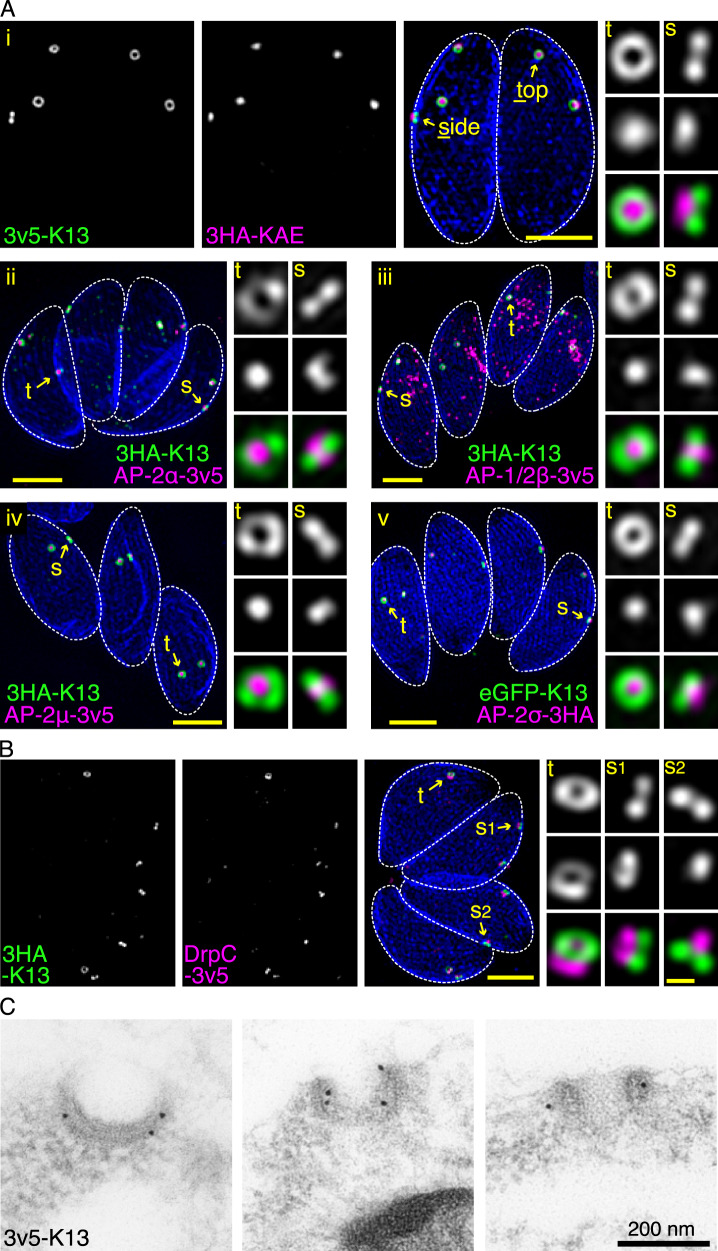
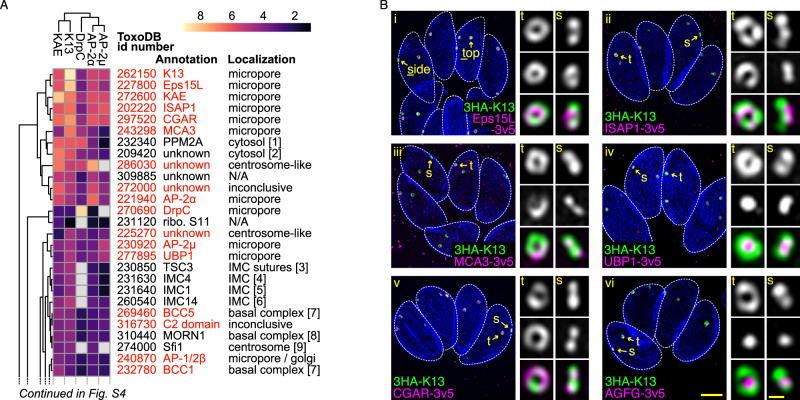
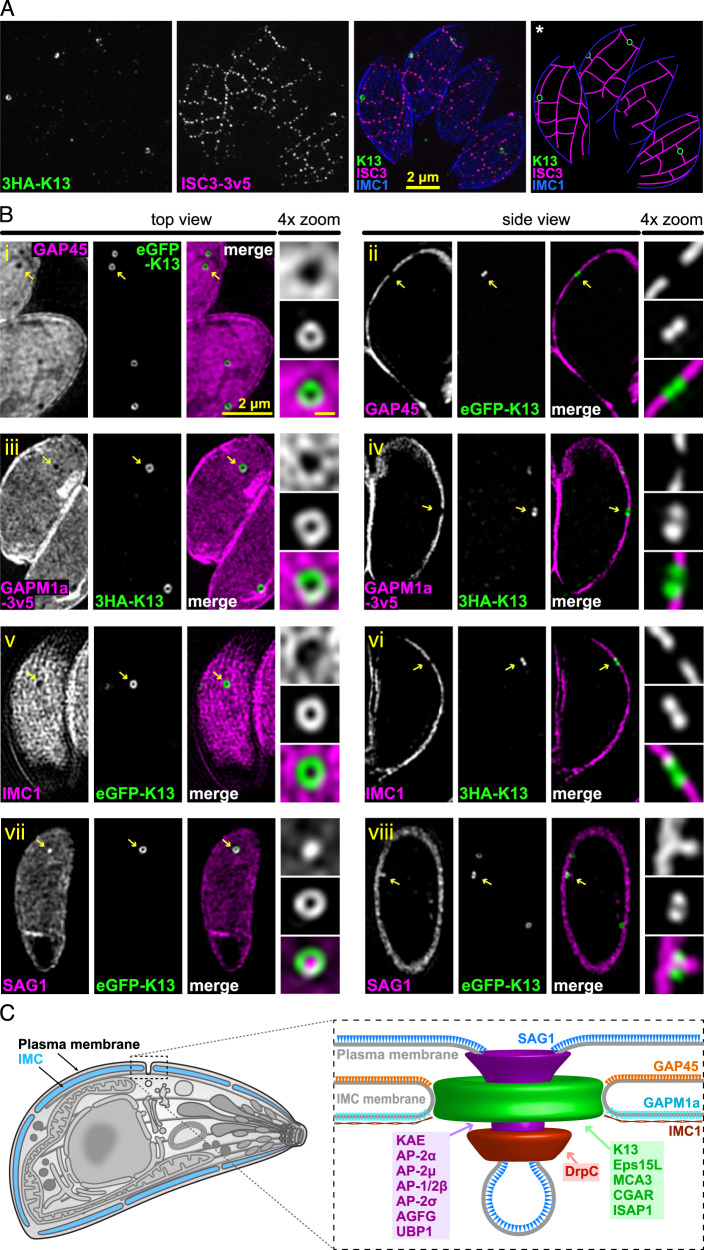
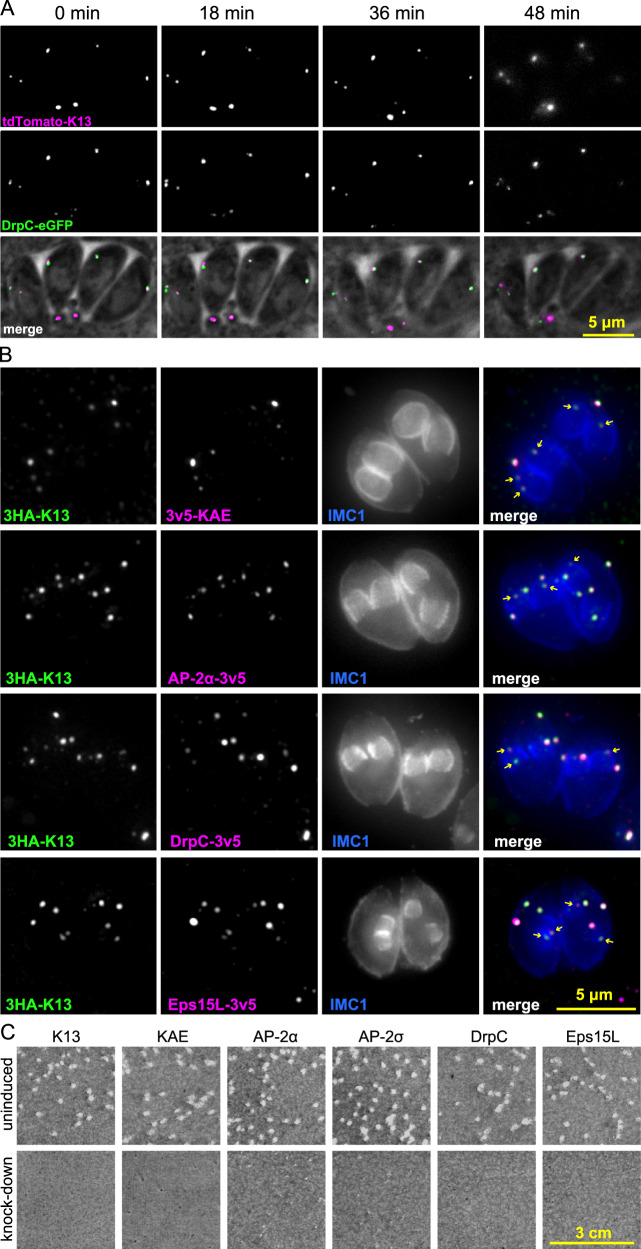
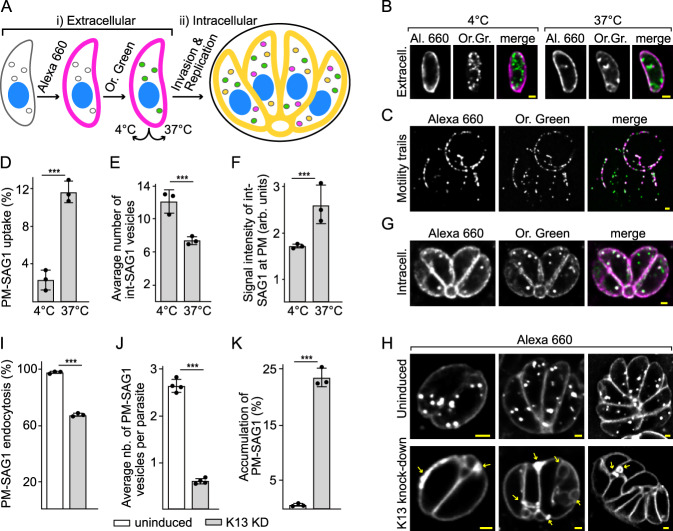
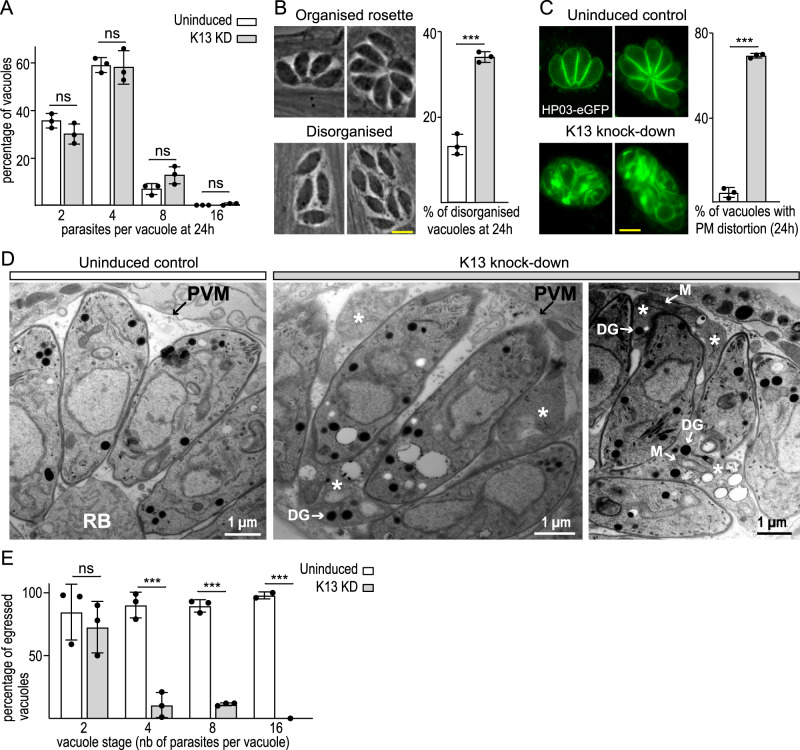
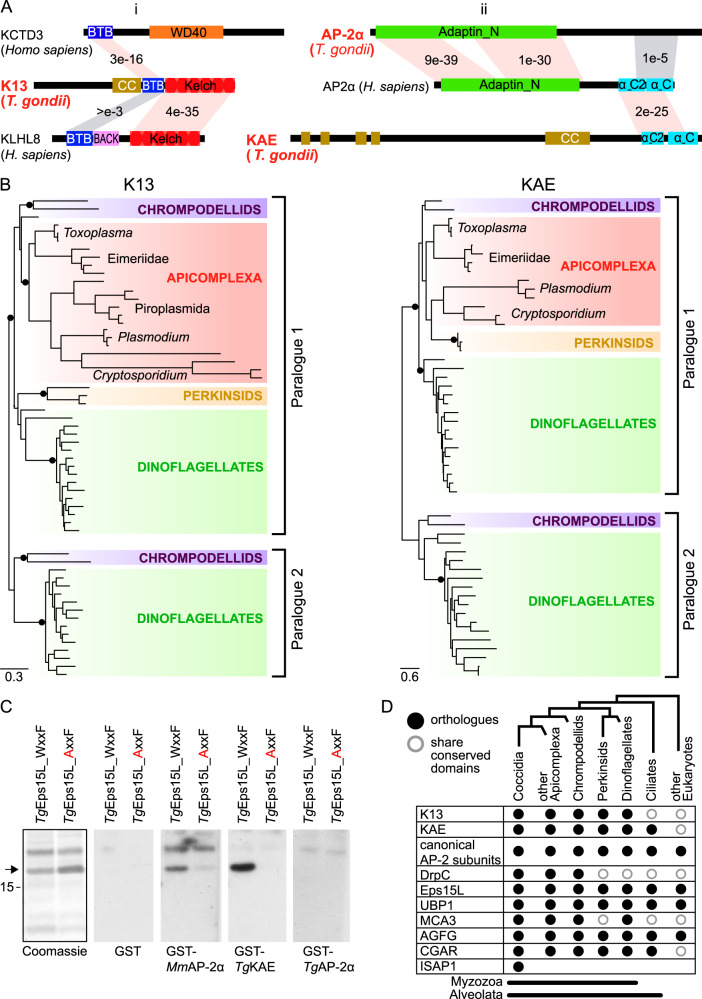
Apicomplexan parasites have immense impacts on humanity, but their basic cellular processes are often poorly understood. Where endocytosis occurs in these cells, how conserved this process is with other eukaryotes, and what the functions of endocytosis are across this phylum are major unanswered questions. Using the apicomplexan model Toxoplasma, we identified the molecular composition and behavior of unusual, fixed endocytic structures. Here, stable complexes of endocytic proteins differ markedly from the dynamic assembly/disassembly of these machineries in other eukaryotes. We identify that these endocytic structures correspond to the 'micropore' that has been observed throughout the Apicomplexa. Moreover, conserved molecular adaptation of this structure is seen in apicomplexans including the kelch-domain protein K13 that is central to malarial drug-resistance. We determine that a dominant function of endocytosis in Toxoplasma is plasma membrane homeostasis, rather than parasite nutrition, and that these specialized endocytic structures originated early in infrakingdom Alveolata likely in response to the complex cell pellicle that defines this medically and ecologically important ancient eukaryotic lineage.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Functional screening reveals Toxoplasma prenylated proteins required for endocytic trafficking and rhoptry protein sorting.mBio. 2023 Aug 31;14(4):e0130923. doi: 10.1128/mbio.01309-23. Epub 2023 Aug 7. mBio. 2023. PMID: 37548452 Free PMC article.
-
The ZIP Code of Vesicle Trafficking in Apicomplexa: SEC1/Munc18 and SNARE Proteins.mBio. 2020 Oct 20;11(5):e02092-20. doi: 10.1128/mBio.02092-20. mBio. 2020. PMID: 33082261 Free PMC article.
-
The Toxoplasma micropore mediates endocytosis for selective nutrient salvage from host cell compartments.Nat Commun. 2023 Feb 22;14(1):977. doi: 10.1038/s41467-023-36571-4. Nat Commun. 2023. PMID: 36813769 Free PMC article.
-
Apicomplexa micropore: history, function, and formation.Trends Parasitol. 2024 May;40(5):416-426. doi: 10.1016/j.pt.2024.03.008. Epub 2024 Apr 17. Trends Parasitol. 2024. PMID: 38637184 Review.
-
Endocytosis in different lifestyles of protozoan parasitism: role in nutrient uptake with special reference to Toxoplasma gondii.Int J Parasitol. 2001 Oct;31(12):1343-53. doi: 10.1016/s0020-7519(01)00252-1. Int J Parasitol. 2001. PMID: 11566302 Review.
Cited by
-
HOPS/CORVET tethering complexes are critical for endocytosis and protein trafficking to invasion related organelles in malaria parasites.PLoS Pathog. 2025 Apr 8;21(4):e1013053. doi: 10.1371/journal.ppat.1013053. eCollection 2025 Apr. PLoS Pathog. 2025. PMID: 40198740 Free PMC article.
-
Characterization of atypical BAR domain-containing proteins coded by Toxoplasma gondii.J Biol Chem. 2024 Dec;300(12):107923. doi: 10.1016/j.jbc.2024.107923. Epub 2024 Oct 24. J Biol Chem. 2024. PMID: 39461477 Free PMC article.
-
Atlas of Plasmodium falciparum intraerythrocytic development using expansion microscopy.Elife. 2023 Dec 18;12:RP88088. doi: 10.7554/eLife.88088. Elife. 2023. PMID: 38108809 Free PMC article.
-
Atlas of Plasmodium falciparum intraerythrocytic development using expansion microscopy.bioRxiv [Preprint]. 2023 Oct 9:2023.03.22.533773. doi: 10.1101/2023.03.22.533773. bioRxiv. 2023. Update in: Elife. 2023 Dec 18;12:RP88088. doi: 10.7554/eLife.88088. PMID: 36993606 Free PMC article. Updated. Preprint.
-
TBC9, an essential TBC-domain protein, regulates early vesicular transport and IMC formation in Toxoplasma gondii.Commun Biol. 2024 May 18;7(1):596. doi: 10.1038/s42003-024-06310-6. Commun Biol. 2024. PMID: 38762629 Free PMC article.
References
-
- Autoridad Nacional del Servicio Civil. World malaria report 2021. Angew. Chem. Int. Ed.6, 951–952 (2021).
-
- Striepen, B. Time to tackle cryptosporidiosis. Nature503, 189–191 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
